|
Г.Э. Фельдман
Джон Бэрдон Сандерсон 1892-1964 Изд. "Наука", Москва, 1976 г.. |
|
|
Г.Э. Фельдман
Джон Бэрдон Сандерсон 1892-1964 Изд. "Наука", Москва, 1976 г.. |
Глава II
Научное творчество
В одном мгновенье видеть вечность,
Огромный мир - в зерне песка,
В единой горсти - бесконечность
И небо - в чашечке цветка.Уильям Блейк
(перевод С. Маршака)
ГенетикаВспоминая о своем первом знакомстве с генетикой, Холдейн писал: "Моя связь с генетикой начинается с 1901 г., т.е. годом спустя после переоткрытия законов Менделя. Мало кто может похвастаться таким внушительным стажем. В 1901 г., когда мне минуло восемь лет, я с отцом присутствовал на дискуссии научного студенческого общества в Оксфорде, где прослушал лекцию Дербишера об открытиях Менделя. Мне трудно было все понять, но это было очень интересно. Лишь в 1911 г. я приступил к научным исследованиям по генетике" *.
* J.B.S. Haldane. Biological possibilities of the human species in the next ten thousand years. - In: "Man and his future". London, 1963, p. 337.Со слов Холдейна известно, что в 1908 г. в библиотеке отца он ознакомился с работой Дербишера, напечатанной в журнале "Биометрика". Он заинтересовался вопросом расположения генов в хромосомах растений и задумался над возможностью обнаружения этого явления у животных. Его сестра Нейоми в эту пору увлекалась разведением морских свинок, и, возможно, поэтому первое исследование Холдейн провел именно на этих животпых.В это время на базе Института садовых культур Джона Иннеса уже была создана школа английских генетиков во главе с Уильямом Бэтсоном (1861-1926), а в Кембриджском университете кафедрой генетики руководил Р. Пеннет (1875-1967).
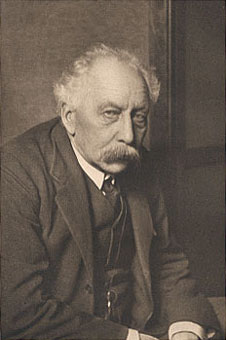
Уильям Бэтсон
Свою первую студенческую работу по генетике под несколько претенциозным названием "Сравнительная морфология зародышевой плазмы" молодой Холдейн доложил в 1911 г. в Оксфорде на студенческом семинаре по представлению своего учителя зоологии Е.С. Гудрича, ученика известного биолога Рея Ланкестера (1847-1929). Ободренный успехом Холдейн обратился к Р. Пеннету с просьбой опубликовать это исследование. Пеннет посоветовал провести дополнительные наблюдения, используя при этом не только морских свинок, но и мышей, что значительно задержало завершение работы, по существу направленной на решение центральной проблемы науки о наследственности - расположения генов в хромосомах.
В 1905-1910 гг. в лаборатории выдающегося американского биолога Эдмунда Вильсона (1856-1939) были осуществлены эксперименты, доказавшие, что именно хромосомы определяют формирование мужского или женского пола у организмов. По представлению Вильсона и его учеников (и в первую очередь Томаса Гента Моргана), среди хромосом любых организмов можно найти так называемые половые хромосомы, сочетаниями которых определяется, быть ли созревающему организму мужским или женским. Авторы полагали, что в этих хромосомах находятся особые факторы, которые и определяют развитие организма в мужскую или женскую сторону.
Начиная с 1910 г. Морган провел серию классических генетических исследований на великолепной экспериментальной модели - дрозофиле. Удобство работы с этим объектом обеспечивалось двумя главными его особенностями - быстротой размножения (одно поколение Dr. melanogaster можно было получить всего за один месяц) и возможностью работать с огромными количествами организмов (десятками, а подчас даже с сотнями тысяч мух в одном эксперименте). Благодаря этому Моргану удалось довольно быстро прийти к заключению, что гены, контролирующие различные признаки, располагаются не как попало, случайно, а в определенном порядке, закономерно. В одной хромосоме можно было обнаружить одни гены, в другой - другие и т. д. Естественно, что гены, находящиеся в одной хромосоме, передавались при скрещивании единой группой, или, как стали говорить генетики, сцепленно. В частности, гены, передаваемые с хромосомами, определяющими тот или иной пол (половыми хромосомами), получили название "сцепленные с полом".
Однако как обстоит дело с наследованием признаков и определяющих их генов у других организмов (у растений и животных), оставалось неясным. Приложимы ли правила сцепления к ним? Какова количественная характеристика сцепления? Сколько генов образуют одну группу сцепления? Множество вопросов было поставлено генетиками, но чаще всего даже подходов к их решению наметить не удавалось.
Организмы, с которыми начал работать молодой Холдейн, нельзя было назвать ни новыми для генетиков, ни традиционно изучавшимися. Кое-что было про них известно. В течение первого десятилетия XX в. было опубликовано 10 исследований по окраске мышей. Уже через два года после вторичного "переоткрытия" законов Менделя Де Фризом, Корренсом и Чермаком во Франции была опубликована работа зоолога Л. Кэно под интригующим названием "Закон Менделя и наследование окраски у мышей", в которой автор проанализировал характер наследования белой и серой окраски у 198 серых и 72 белых мышей. Хотя экспериментальный материал был невелик, автору удалось отметить, что наследование подчиняется менделевскому моногибридному характеру. Этой теме Л. Кэно в период 1903-1908 гг. посвятил еще пять публикаций.
Вслед за ним исследования по окраске мышей выполнили Аллен (1904), Дербишер (1903, 1904), ученица и сотрудница Бэтсона мисс Дёрэм (1908, 1911). Однако никому из них не приходило в голову попытаться изучить сцепление этих признаков. Дело было даже не в том, что для такой работы требовался огромный экспериментальный материал, но и в отсутствии должного математического таланта. Но именно математический склад ума был сильной стороной Холдейна. По-видимому, у Холдейна был и еще один моральный стимул взяться за разрешение этой проблемы. Дело в том, что его учитель Бэтсон в этот период ожесточенно оспаривал выводы школы Моргана о сцеплении генов. Им были поставлены весьма тонко разработанные эксперименты, из которых следовало, что законы Менделя лишены универсальности. Бэтсон потратил много лет на полемику с приверженцами менделизма и обнаружил наиболее яркие примеры, противоречащие его учению. Скепсис и недоверие к общепринятому в науке были одной из наиболее характерных черт этого большого ученого. По-видимому, Холдейн унаследовал эти особенности научного и житейского характера своего наставника, пронеся их через всю свою долгую жизнь. Он также никогда и ничего не принимал на веру. И если сегодня имя Бэтсона вспоминается не иначе как в связи с подтверждением законов Менделя и открытий Моргана, то именно потому, что углубленное изучение "исключительных случаев" с новой силой подтвердило обоснованность этих революционных открытий.
В своем очерке памяти Бэтсона Холдейн писал:
"В течение восьми лет Бэтсон яростно нападал на теорию Моргана. И не потому, что считал ее по сути своей неверной, но потому что был убежден, что она не подтверждена наблюдениями, а также потому что склад его ума и его обширные познания из истории науки воспитали в нем недоверие к пространным рассуждениям, какими бы убедительными они ни были. Когда представилась возможность увидеть картину деления, Бэтсон отправился в Америку и вернулся приверженцем теории Моргана, хотя и с некоторыми оговорками и внутренней сдержанностью, которые, я полагаю, в будущем будут в значительной степени оправданы" *.Возможно, что пониманию и восприятию Бэтсоном законов Моргана в немалой степени способствовали результаты, полученные молодым Джоном Холдейном, работавшим в эти годы в его лаборатории и у Р. Пеннета *. Холдейну с соавторами удалось убедительно показать сцепление наследственных признаков у мышей. Мировая война прервала занятия в лаборатории. Холдейн додумывал выводы и дописывал работу уже в окопах. По представлению Бэтсона это исследование было опубликовано в 1915 г. В нем авторы проанализировали данные Дербишера (1904) относительно характера наследования розовой окраски глаз и альбинизма у мышей и подвергли их тщательной проверке. В результате было установлено наличие сцепления генов окраски волос С и темной окраски глаз Е. Позднее явление сцепления у грызунов, в частности у крыс, было обнаружено и другими исследователями.* J.B.S. Haldane. Possible worlds and other essays. London, 1927, p. 137.* Во время работы у Бэтсона и Пеннета Холдейн впервые встретился с Н.И. Вавиловым, дружбой с которым он гордился до конца своих дней.С другой сторопы, научная страстпость, желание и умение работать только на переднем крае, а не "доделывать зады" была заложена в Холдейне не без участия Бэтсона, который "никогда не скрывал своего презрения к второсортному исследованию или второсортным мыслям. В поисках истины он был беспощаден и безжалостен к мнению других, так же как к собственному. Его путь в науку лежал через морфологию, и все его работы отмечены печатью мыслителя-морфолога. Порой непоправимый скептицизм Бэтсона можно объяснить именно тем, что ему были чужды рассуждения, далекие от морфологического образа мышления. Не было случая, чтобы он, если мне в ходе спора удавалось убедительно доказать ошибочность его доводов, не признал свою неправоту. При этом он никогда не упрекал меня, что я несу вздор, хотя зачастую бывало и так" *. Эта характеристика в полной мере приложима и к самому Холдейну.* J.B.S. Haldane. Possible worlds and other essays, p. 137.Следует отметить, что вопрос о сцеплении генов оставался в центре интересов ученого до конца его дней и изучался им на разных объектах (животных и растениях) в соответствии с новыми успехами в отдельных разделах науки о наследственности.В одной из работ 1919 г. Холдейном была предложена математическая модель кроссинговера и сделаны соответствующие выводы. Все его расчеты были проверены и подтверждены биологическими экспериментами. Предложенный им метод позволял учитывать возможные погрешности и оказался наиболее точным для того времени, а формула Холдейна до сих пор используется в опытах по изучению рекомбинации генов.
В следующем сообщении того же 1919 г. им была разработана формула определения соотношения между расстоянием на карте хромосом и частотой рекомбинаций ("картирующая функция Холдейна"). В работе 1920 г. Холдейн обратил внимание на наличие сцепления двух факторов окраски у американской саранчи (Paratettix taxanus) при изучении данных Наборса, а в 1921 г. впервые подметил случай частичного сцепления признаков у кур. Уже в эту пору проявился удивительный талант Холдейна-синтетика, внимательно следящего за развитием всех направлений науки о наследственности и умеющего обобщать данные, поступающие из разных источников. Из ранних работ Холдейна явствует, что сильнейшей стороной его научной деятельности является блестящее владение математическим аппаратом при анализе любых данных: сцепление генов, нарушение сцепления из-за обмена участками генов - кроссинговера - между хромосомами. Выше уже упоминалось, что формула кроссинговера, выведенная Холдейном, до настоящего дня используется в качестве основного математического аппарата для описания вероятностного обмена генами на определенном участке хромосом. Этот вопрос всегда оставался в центре его внимания. Он последовательно усовершенствовал математическую разработку этого важнейшего вопроса (1944, 1946, 1947) в поисках биологических закономерностей кроссинговера у различных организмов: гороха (1930), львиного зева (1929, 1933), комаров (1946, 1947).
Начиная с 1930 г. Холдейн вплотную занялся изучением наследственности у человека. В работе 1934 г. "Относительная эффективность двух методов измерения сцепления генов у человека" он весьма критически отнесся к статистическому материалу, полученному другими авторами, подвергнув его проверке методом математического анализа. Сравнивая частоту мутаций у дрозофилы и человека, Холдейн на основании математических расчетов приходит к выводу, что человек более мутабилен, т.е. подвержен мутационным изменениям, чем дрозофила *.
* J.B.S. Haldane. Mutation rates in man, - "Nature", 1935, v. 135, p. 907.Другой важнейшей стороной исследований Холдейна было то, что, занимаясь отвлеченными математическими аспектами теории кроссинговера, он никогда не забывал о физиологической целостности организма, о влиянии обмена веществ, возраста, числа хромосом на проявление генной активности. В работе 1925 г. совместно с Ф.Е. Крoу им было показано влияние возраста организма (опыты на курах) на частоту процесса обмена генов. Через несколько лет, в 1933 г., русский цитолог и генетик М.С. Навашин (1905-1973) при изучении наследственности у растений описал роль длительности хранения семян до их высева на частоту мутационного процесса. Обе эти работы - Холдейна и Навашина - положили начало важнейшей серии исследований в области физиологической генетики.Первоначальные эксперименты и сделанные на их основе обобщения касались только тех организмов, которые несли нормальное число пар хромосом. Но еще в конце прошлого века (1890-1904) русский биолог И.И. Герасимов (1867-1920) описал явление, впоследствии названное полиплоидией, когда число хромосом увеличивается в несколько раз. Сформулированная Морганом и его учениками и последователями теория кроссинговера касалась лишь перекреста генов у нормальных (диплоидных) организмов. Вопрос же о приложимости выводов этой теории к полиплоидам оставался неисследованным.
В серии сообщений, подготовленных совместно с Де Уинтом (1931, 1933, 1935), Холдейн исследовал проблему сцепления генов у диплоидных и тетраплоидных растений (на примере примулы) на основе разработанной им довольно сложной математической теории определения сцепления в случае полиплоидии (1930). Ему удалось показать, что процент кроссинговеров у диплоидных растений близок к таковому у тетраплоидных.
Авторы исследовали локализацию 15 факторов в четырех хромосомах (из 12) у диплоидных растений, а также изучили случаи сцепления у тетраплоидной примулы. Ими описаны шесть типов сцепления, наблюдаемых в случае трех пар факторов у той же примулы. В работе подчеркивается, что у тетраплоидных растений кроссинговер протекал одновременно только лишь между двумя гомологичными хромосомами, хотя их было в клетке четыре. После кроссинговера эти хромосомы расходились к разным полюсам. При этом отмечается, что данные, полученные на примуле, необходимо проверить на других видах тетраплоидных растений.
Удивительная способность к обобщению разрозненных наблюдений и к формулированию важнейших закономерностей сказалась в работе Холдейна "Соотношение полов и стерильность одного пола у гибридных животных" (1922). В ней был тщательно обработай весь доступный к тому времени материал по вопросу о потомстве межвидовых гибридов первого поколения. Сформулированное им положение, известное под названием "правило Холдейна", в значительной степени не утратило своего значения в настоящее время.
Согласно генетической теории образования мужского и женского пола (Вильсон, Морган) разпые полы определяются различным сочетанием хромосом. Если обе формирующие зародыш половые клетки (их называют также гаметами) - сперматозоид и яйцеклетка - несут одинаковые половые хромосомы (например, две Х-хромосомы), то образовавшийся организм называют гомогаметным. В том случае, когда одна гамета привносит в зародыш хромосому одного типа (Х-хромосому), а другая гамета хромосому другого типа (Y-хромосому), то возникает гетерогаметный организм. Но чтобы узнать, к какому типу относится данный организм, необходимо провести серию трудоемких генетических и цитологических экспериментов. Правило Холдейна, в конечном счете, было направлено на решение именно этой проблемы. Оно гласит: если в потомстве межвидовых гибридов один из полов встречается реже, полностью отсутствует или стерилен, то этот пол является обычно гетерогаметным. Нежизнеспособность или стерильность этого гетерогаметного пола может быть результатом взаимодействия между Y-хромосомой одного вида и Х-хромосомой, аутосомами или цитоплазмой другого вида. При наличии механизма определения пола типа ХО гибель, пониженная жизнеспособность или стерильность межвидовых гибридов могут быть следствием взаимодействия аутосом или цитоплазмы одного вида с единственной Х-хромосомой, полученной от другого скрещиваемого вида.
В 1922 г. Н.И. Вавилов сформулировал закон гомологических рядов наследственной изменчивости. Он пришел к выводу, что близкородственные по происхождению виды растений и животных имеют сходные ряды наследственной изменчивости. Эта гомологичность обусловлена родственным происхождением организмов, близостью их генетического аппарата. * * *
Анализу гомологических рядов наследственной изменчивости посвящена работа Холдейна "Сравнительное исследование окраски у грызунов и хищников" (1927). Рассматривая вопросы генетической гомологии на примерах двух групп животных, Холдейн обобщил типы обнаруженных мутаций в пределах изученных отрядов млекопитающих, связав свои наблюдения с эволюционной теорией.
Мысль Холдейна о применении принципов гомологии к генам не утратила своей актуальности, хотя с тех пор прошло почти полвека. Тогда Холдейн отмечал, что "структуры в двух видах животных являются гомологичными при условии, что они соответствуют одной и той же структуре у общего предка". И при этом подчеркивал, что абсолютного критерия для гомологии не существует и нет уверенности в том, что разные формы генов ведут свое происхождение от гена общего предка. Можно предположить, что в отдельных случаях гены могут иметь совершенно самостоятельное происхождение. Со свойственной ему дальновидностью Холдейн ставит вопрос о возможности более близкой связи между генами, чем гомология, а именно - об их химической идентичности. На основании данных о сцеплении он делает допущение, что гены по своим размерам одного порядка с белковыми молекулами.
Рассматривая вопрос о пластичности наследственности, о способности видов к наследственной изменчивости, Холдейн отмечал, что вид может оказаться весьма подверженным изменчивости в одном отношении и весьма мало изменчивым в другом. Он полагал, что нужно с большой степенью осторожности судить о большей или меньшей пластичности вида. Позднее по этому поводу Холдейн писал: "До тех пор пока этот вопрос (генетическая пластичность) не будет изучен в плане сравнительной генетики, генетика лишь в малой степени сможет способствовать познанию основных направлений эволюционного процесса" *.
* J.B.S. Haldane. The theory of evolution before and after Bateson. - "J. of Genetics", 1958, v. 56, p. 12.В настоящее время в связи с достижениями молекулярной генетики разработка нового направления - сравнительной генетики - представляется особенно перспективной, если при этом принять во внимание успехи структурного исследования некоторых белков - таких, к примеру, как гемоглобин и цитохром-С, которые являются прямыми продуктами действия генов.
Популяционная генетика и эволюцияЕще в середине прошлого века Дарвин установил основные факторы эволюции: наследственность, изменчивость и естественный отбор.
Естественный отбор, т.е. выживание наиболее приспособленных форм животных и растений, был обоснован Дарвином с достаточной очевидностью. Что же касается наследственности и изменчивости, то здесь положение было не совсем яспым в силу недостаточности знаний о механизмах наследственности. Дарвин считал, что отбору могут подвергаться как случайные изменения, так и изменения, вызванные упражнением органов. Вслед за общепризнанным в те времена взглядом он верил в так называемую слитную (blending) наследственность - предполагалось, что признаки родителей смешиваются у потомков подобно воде с чернилами; дискретность наследственных факторов еще не была известна. Поэтому Дарвип недостаточно осветил вопрос о причинах, определяющих возникновение наиболее приспособленных форм. Он и не мог этого сделать, поскольку корпускулярная природа наследственности еще не была обоснована. А работы Менделя хотя и были опубликованы, Дарвин их не знал.
Положение, сложившееся к тому времени в науке, дало повод противникам теории Дарвина отрицать роль естественного отбора как фактора эволюции. Например, английский инженер Ф. Дженкин писал, что любой новый признак в результате скрещивания будет постепенно разбавляться от поколения к поколению, пока не исчезнет совсем. При такой ситуации естественный отбор не может играть, конечно, никакой роли, как бы велико ни было первоначальное изменение в организме. Теория Дженкина (1867) о разбавлении из поколения в поколение любого ценного признака при скрещиваниях мутанта с окружающими его исходными формами до постепенного его исчезновения из популяции просуществовала до конца XIX в. и отпала только после возникновения экспериментальной генетики, основанной на законах Менделя. В доменделевские времена никто не был в состоянии опровергнуть тезис Дженкина о нивелирующем результаты отбора действии скрещиваний. Этот тезис удерживался в науке вплоть до середины 20-х годов XX в. и окончательно отпал только после того, как советский генетик С.С. Четвериков (1880-1959) связал между собой эволюционный принцип Дарвина и менделевские законы наследственности *.
* С.С. Четвериков. О некоторых моментах эволюционного учения с точки зрения современной генетики. - "Журнал экспериментальной биологии", 1926, т. 2, вып. 1, стр. 3.Используя математическую формулу свободного скрещивания Харди-Вайнберга (1908), Четвериков сумел доказать, что вновь возникшие мутации не могут исчезнуть из популяции в ходе скрещивания, и показал значение отбора в этих условиях. Кстати сказать, Холдейн хорошо знал и высоко ценил эту замечательную работу; в памяти участников V Международного конгресса генетиков, проходившего в Берлине в 1927 г., остался характерный эпизод: по окончании сообщения, сделанного С.С. Четвериковым, к трибуне бросился Холдейн, заключил докладчика в объятия и горячо расцеловал.Таким образом, в середине 20-х годов перед исследователями возник целый ряд проблем, которые не всегда можно было решить экспериментально. Поэтому математический метод, одним из создателей которого был Холдейн, стал мощным орудием в исследовании проблем генетики популяций и эволюции.
Большая серия работ Холдейна посвящена математической теории естественного отбора. К этому времепи действие естественного отбора на состояние популяций было выяснено только для простейших случаев. Сущность проблемы, которую предстояло разрешить, была кратко сформулирована Холдейном в первом абзаце первого сообщения, опубликованного им в 1924 г. в "Ученых записках философского общества Кембриджского университета", следующим образом: "Естественный отбор должен быть охарактеризован прежде всего с количественной стороны. Для того чтобы утверждать, что естественный отбор в состоянии объяснить известные факты эволюции, мы должны показать, что он может не только заставить вид изменяться, но и воздействовать на темп мутаций в настоящем и прошлом".
В своем первом сообщении Холдейн подчеркивает тот факт, что элементарной единицей эволюции является не особь, а популяция. Он разбирает различные состояния популяций, для которых дает общие характеристики, изучает такие важные для эволюции факторы, как избирательное спаривание и действие близкородственного скрещивания (инбридинга) в связи с отбором, исследует полигенные системы, в некоторых случаях включающие и взаимодействия между генами, отбор в зависимости от частоты встречаемости генов в популяции, отбор у полиплоидов, последствия перекрывания одного поколения другим в противоположность дискретности поколений, влияние частичной изоляции.
В 1924 г. Холдейн разработал таблицу, при помощи которой можно было без труда определить изменения частоты генотипов и интенсивность отбора во времени.
Основные исследования 1924-1934 гг. были посвящены систематическому изучению темпа изменения популяции под влиянием отбора и мутаций. Холдейн постулировал, что элементарным процессом эволюции является увеличение или уменьшение концентрации отдельных генов в популяции. При помощи математического моделирования он показал, что можно рассчитать, как изменяется относительная частота встречаемости отдельных генов при заданной интенсивности отбора - быстро или медленно. Количественные расчеты позволили Ходдейну нарисовать общую картину темпа эволюционного процесса. Ему удалось показать, что медленный темп мутирования, характерный для большинства генов, по-видимому, не является лимитирующим фактором в эволюции.
Математический анализ процессов эволюции, проведенный Холдейном, представляет собой дальнейшее развитие и конкретизацию учения Дарвина о естественном отборе с учетом новейших достижений генетики. Он отвечал интересам и нуждам животноводов и растениеводов, занимающихся выведением новых пород на основании теории отбора.
В этой серии исследований Холдейн выступил как пионер нового направления - математической генетики, исследуя изменение частоты встречаемости генов в популяциях в ходе естественного и искусственного отбора. Он подверг анализу вопросы динамики, эволюции, разработал уравнения, отражающие темп изменения состава популяций для случаев слабого, интенсивного отбора и отбора средней интенсивности. При этом Холдейн полагал, что коэффициент отбора, как правило, является постоянным. В некоторых случаях он может быть переменной величиной по отношению к отдельно взятому гену, или в отдельных случаях по отношению к отдельно взятому генотипу с учетом двух или большего числа взаимодействующих локусов.
Одновременно с С.С. Четвериковым и Дж. Холдейном над математической интерпретацией эволюционной микроизменчивости растительных и животных организмов работали еще два крупных исследователя: Р. Фишер в Англии и С. Райт в США. Всех их объединял дух дружественного соревнования, направленного на развитие теорий Дарвина и Менделя в свете успехов генетики 20-30-х годов; Р. Фишер в своей теории основное значение придавал благоприятным мутациям, которые достигают частоты, предохраняющей их от случайной утраты. Он считал, что повышение приспособленности любого организма в любое время зависит от генотипической варианты в отношении приспособленности. Под генотипической вариантой он подразумевал ее аддитивный компонент *.
* См. R.A. Fisher. The general theory of natural selection. Oxford, Clarendon Press, 1930.

Рональд Фишер

Сьюэл Райт
С. Райт полагал *, что в отношении частоты генов имеет место локальная дифференциация. Тем самым он отбросил общепринятое в популяционной генетике допущение о вполне случайной скрещиваемости особей внутри отдельной популяции. В таком случае может образоваться несколько пиков приспособленности в противоположность одному основному пику в гомогенных видах. Подобное положение может установиться вследствие различных процессов, которые являются случайными по отношению к эволюции вида в целом. Райт постулировал три фазы дифференцировки внутри популяции: фаза случайного дрейфа систем генов от равномерности их распределения, фаза локального массового отбора по отношению к новой системе концентраций генов, фаза избыточного роста популяции и отбора по системам генотипов в целом (межгрупповой отбор), ведущего к постепенному преобразованию всего вида.
* Cм. S. Wright. Evolution in a Mendelian population. - "Genet.", 1931, v. 16, p. 97.Холдейн неоднократно подчеркивал, что эволюция не может поддерживаться каким-либо одним процессом. Межгрупповой отбор не будет играть никакой роли, если нет достаточной изоляции между группами внутри вида. В гомогенных популяциях при постоянных условиях в течение длительного периода процесс эволюции протекает очень медленно и определяется только частотой возникновения благоприятных мутаций. При постепенном изменении условий эволюционный процесс может ускориться за счет накопления мутаций, которые до этого были частично неблагоприятными, но в новых условиях становятся благоприятными.Оптимальная ситуация для значительного эволюционного сдвига будет, очевидно, связана с определенным сочетанием подходящей генетической структуры популяции и длительности периода изменения условий существования.
Холдейн математически определил, что и межгрупповой отбор в некоторых случаях очень эффективен. Это касается, например, популяций человека, по крайней мере, в период резкого увеличения численности *. Он описал условия, при которых мутация может сохраниться в популяции, вопреки действию отбора, и пришел к выводу, что только отбор пли только мутации не могут целиком определить конечный эффект **.
* J.B.S. Haldane. The rate of mutation of human genes. - "Proc. 8th Intern. Congr. of Genetics. Hereditas Suppl.", 1949, p. 267.Допустим, р обозначает частоту мутирования гена за одно поколение (обычно р = 10-6), а k - коэффициент отбора, причем к больше, чем р (речь идет о неблагоприятных мутациях). В таком случае равновесие достигается тогда, когда отношение неблагоприятных фенотипов к благоприятным равно р/k при рецессивной мутации и 2р/k - при доминантной в свободно скрещивающейся популяции. Поэтому, если величина к не настолько мала, чтобы быть одного порядка с р, новая мутация не распространится в значительной степени. Даже при высокой частоте мутирования отрицательный отбор при значениях к не менее 1/2000 делал бы редкой любую рецессивную мутацию. Таким образом, чтобы вызвать большие изменения в видах, частота мутирования должна превышать существующую в сотни и тысячи раз. Однако один отбор вряд ли играет существенную роль. Ни один из этих процессов в отдельности не может служить основой для продолжительной эволюции. Материал, с которым имеет дело отбор, должен поставляться мутациями.** J.B.S. Haldane. The rate of spontaneous mutation of a human gene. - "J. of Genetics", 1935, v. 31, p. 317.
Много внимания Холдейн уделял такому существенному вопросу, как эволюция доминантности *. К этому времени существовало две точки зрения. Фишер полагал, что доминантность эволюционирует посредством подбора генов-модификаторов, действие которых направлено на ликвидацию вредных для организма последствий мутации. Так как мутация в гомозиготном состоянии часто оказывается особенно неблагоприятной, то и модифицированию в основном подвергаются более многочисленные гетерозиготы. Райт, напротив, считал, что отбираются не модификаторы, а наиболее активные по сравнению с исходным аллели **.
* См. J.B.S. Haldane. A note of Fisher's theory of the origin of dominance and on the correlation between dominance and linkage. - "Amer. Naturalist", 1930, v. 64, p. 87; J.B.S. Haldane. The theory of the evolution of dominance. - "J. of Genetics", 1939, v. 37, p. 365.Холдейн рассмотрел три причины доминантности и показал возможное распространение этих явлений в природе *. Причины эти следующие: 1) фишеровский эффект (в редких случаях); 2) отбор наиболее активного мутантного гена по сравпению с исходным (в большинстве случаев - толкование Райта) - и, наконец, 3) дупликации, составляющие небольшую группу случаев.** См. S. Wright. Evolution of a mendelian population. - "Anatomical Record", 1929, p. 287.
* J.B.S. Haldane. The theory of the evolution of dominance. - "J. of Genetics", 1939, v. 37, p. 365.В больших аутбредных популяциях отбор будет увеличивать концентрацию генов, модифицирующих жизнеспособность гетерозигот, в направлении, приближающем их к норме. Гомозиготы модифицируются уже после гетерозигот. В таких популяциях фишеровский эффект наиболее распространен. В аутбредных, но небольших популяциях наблюдалась бы та же картина, но гомозиготы подвергались бы модификации в большей степени.В инбредных популяциях отбор модификаторов для гетерозигот должен протекать очень медленно, а для гомозигот и вовсе отсутствовать. Если же есть хотя бы небольшой процент аутбридинга (порядка 1%), отбор гомозигот будет значительным, особенно после того, как они достигнут приспособленности 1 - с/2 (с - процент аутбридинга). Такого рода популяции распространены среди растений (например, Pisum и Lathyrus).
Касаясь физиологических основ доминирования, Холдейн исходил из того, что мутантные гены часто оказываются полностью или частично неактивными. В то же время ген дикого типа по фенотипическому проявлению имеет так называемый фактор безопасности (запас прочности). Этот фактор может быть достигнут либо отбором более активного аллеля, либо с помощью отбора модификаторов, направленных на повышение эффективности действия нормального гена.
Накопление модификаторов влечет за собой значительные изменения в организме. Необходимо начинать выпавший процесс снова, если ген неактивен, или ускорить его в 10-100 раз, если ген слабоактивен. С этой точки зрения доминирование проще достигается отбором более активного аллеля.
В 1937 г. в работе "Влияние изменчивости на приспособленность" Холдейн сформулировал понятие "утраты приспособленности", согласно которому влияние повторно возникающих вредных мутаций на величину приспособленности популяции определяется не столько вредоносностью каждой из этих мутаций, сколько частотой ее повторного возникновения в данной популяции. К такому же выводу спустя несколько лет пришел независимо от Холдейна также и Г.Дж. Меллер. В дальнейшем это понятие привело к интенсивному исследованию полиморфных популяций человека и концепции "генетического груза", разработанной Кроу *.
* См. J.F. Crow. The theory of genetic loads. - "Proc. XI Intern. Congress Genetics", 1963, v. 3.Стремление к всестороннему изучению всех количественных аспектов, связанных с интенсивностью естественного отбора, натолкнуло Холдейна на мысль о проведении специального исследования, посвященного клиналь-ной изменчивости, т.е. постепенному пространственному изменению частоты фенотипов *. Изучив распределение признаков (рост, окраска и др.) у оленьей мыши, он предположил, что клинальная изменчивость появляется по той причине, что в одних условиях одна из форм имеет селективное преимущество по сравнению с другой, тогда как в соседней области картина противоположная. Он разобрал взаимодействия между интенсивностью отбора, средним расстоянием расселения за генерацию и скоростью изменения частоты определенного фенотипа в ареале. В данном случае коэффициент отбора был равен приблизительно 0,1%.* Cм. J.B.S. Haldane. The theory of a cline. - "J. of Genetics". 1948, v. 48, p. 277.В работе 1949 г. "Инфекционные болезни и эволюция" Холдейн коснулся еще одного малоразработанного до настоящего времени вопроса - влияния эволюции на распространение инфекционных болезней *. Мысли, высказанные им в этой небольшой по объему, но весьма важной работе о роли естественного отбора в эволюции, о значении сопротивляемости организмов к инфекционным болезням оказали весьма плодотворное воздействие на понимание генетических механизмов и химизма устойчивости растений, животных и человека к инфекционным поражениям. С присущей ему широтой Холдейн поставил вопрос о значении болезней, в том числе и инфекционных, как элиминирующего фактора в процессе естественного отбора, наряду с такими, как борьба с силами природы, голодом, хищниками и внутривидовая борьба. Высказанные им положения имеют общебиологическое значение и целиком приложимы к человеческим популяциям. Холдейн подчеркивал важное значение возникшей в процессе эволюции в ходе естественного отбора устойчивости к болезням и даже предсказывает, что гетерозиготы по гену серповидно-клеточной анемии окажутся невосприимчивыми к малярии, что и было подтверждено в 1954 г. А.С. Эллисоном **. По этому поводу М.Дж.Д. Уайт заметил: "Вполне возможно, что если бы не предсказания Холдейна, то установление зависимости между талассемией и аномальными гемоглобинами, с одной стороны, и устойчивостью к малярии, с другой, было бы отсрочено еще на одно пли два десятилетия" ***.* См. J.B.S. Haldane. Disease and evolution. - "La ricerca scientifica", 1949, v. 19, Suppl., p. 68.Вклад Холдейна в эволюционпую теорию и популяционную генетику, особенно его монография "Факторы эволюции", был по достоинству оценен как его современниками, так и последующим поколением исследователей, работающих в этой области. Основоположник хромосомной теории наследственности Т.Г. Морган писал:** См. В. Маккьюсик. Генетика человека. М., "Мир", 1967, с. 21.
*** М.G.D. White. - "Science", 1969, p. 678.
"Ценным вкладом является интересное обсуждение эволюционной теории, главным образом с генетической точки зрения, в книге Дж.Б.С. Холдейна «Факторы эволюции». Очерк математической теории естественного отбора, данный в этой книге в виде приложения, представляет четкий анализ этой проблемы. Вместе с книгой Р. Фишера «Генетическая теория естественного отбора», представляющей собой более специальный разбор связи между генетикой и естественным отбором, эта работа настолько сдвигает проблему со старого проторенного пути обобщений последних 50 лет в области математических вычислений, что в будущем исследователи эволюции смогут так же специализироваться в этой отрасли математики, связанной с теорией вероятности, как и разрабатывать новые экспериментальные методы для выяснения причин мутаций и действительной роли конкуренции при различных случаях выживания наиболее приспособленных" *.Также высоко заслуги Холдейна-генетика оценивались и через 30 лет. Вот что писал по этому поводу английский генетик Дж.М. Смит в ноябре 1967 г.:* Т.Г. Морган. Экспериментальные основы эволюции. Биомедгиз, 1936."Потребовались огромные усилия Дж. Холдейна, Р. Фишера и С. Райта, чтобы доказать, что подлинная теория эволюции не нуждается в выборе между теориями Дарвина и Менделя - необходимо лишь их объединение в единое целое. Мне думается, что слияние научного творчества Дарвина и Менделя в целостную теорию эволюции является одним из двух или трех действительно важнейших вкладов в биологическую науку нашего столетия" *.Японский генетик М. Кимура в 1966 г. дает следующую оценку работе Холдейна:* J. Maynard Smith. The Listner, 1967, Nov. 2, p. 568."Работы периода создания монографии "Факторы эволюции" можно рассматривать как отправной пункт для дальнейшего развития теории популяций и эволюционной генетики. И особенно печально, что в последнее время среди биологов США, занимающихся вопросами эволюции, появилась тенденция принизить значение этих классических работ, незаслуженно назвав их "генетикой горохового стручка", не предложив при этом никакого эквивалентного подхода к решению вопроса.В последней работе, названной Холдейном "В защиту генетики горохового стручка", - его лебединой песне, подводящей итоги многолетним исследованиям в области популяционной генетики, - он писал:По мере того как математическая подготовка охватывает все более широкие круги молодых биологов, следует выразить надежду, что классические работы Холдейна будут привлекать к себе все более пристальное внимание специалистов" *.
* Цит. по: N.W. Pirie. J.B.S. Haldane. Biographical Memoirs of Fellows of the Royal Society. London, 1966, p. 299.
"Я стремился построить математическую теорию естественного отбора. Мне удалось вычислить равновесие между мутациями различных типов генов и отбором, направленным против мутирования. Как только эта часть работы была завершена, мне представилась возможность в 1932 г. установить частоту мутаций генов у человека. В дальнейшем я внес поправки в свои вычисления, а впоследствии и другие исследователи внесли свои. Определение частоты мутирования генов у человека явилось лишь побочным продуктом моих математических вычислений, по со временем приобрело некоторое политическое значение. Если бы я свою жизнь всецело посвятил научным исследованиям и пропаганде в этой области, а не расширению границ человеческого познания, то я, несомненно, стал бы всемирно признанным «экспертом» в этом вопросе...Потомки, возможно, воздадут должное, а может быть, и не придадут серьезного значения моей теории о «цене естественного отбора», согласно которой эволюционный сдвиг определяется численным выражением генетической смертности. Мне думается, что введенное мною понятие характеризует один из факторов, может быть, наиболее решающий, позволяющий судить о темпе эволюционного процесса. Одни ученые одобрительно отнеслись к моей концепции, другие - критически. Если она окажется ошибочной, то надо думать, что и сама ее порочность прольет некоторый свет на истину или будет способствовать предельному приближению к ней. Количество таких примеров можно приумножить.
Если бы меня судили за то, что я понапрасну растратил свою жизнь, то защитник получил бы дополнительное время по моему делу, если бы оно и было проиграно. В оправдание могу сказать, что мною написано свыше 90 работ по одной лишь генетике горохового стручка и в 50 из них можно найти некоторые оригинальные мысли, не принимая во внимание их значимость или истинность, не говоря о 200 других исследованиях...
Ныне существующие теории по популяционной генетике, несомненно, подвергнутся дальнейшим упрощениям и систематизации. Некоторые из них по своему удельному весу мало чем будут выгодно отличаться от динамической теории XIX в. Это не значит, что эти теории были лишь бесплодной игрой в области алгебраических построений. Приходится испытать много средств и возможностей для достижения даже частичной истины. Однако на этом пути встречается затруднение, состоящее в том, что в результате математического исследования возможная причина явления рассматривается как единственно возможная. .. Может быть, это и не часто случается, но бывает и так. Лучший способ для избежания таких непредвиденных случайностей - это проверка и математический анализ выводов, вытекающих из ряда гипотез, которые на первый взгляд могут показаться весьма искусственными построениями. Нужно всмотреться в природу или материалы, добытые в лаборатории, сопоставить результаты наблюдений с теоретическими заключениями и установить, насколько они истинны или ложны" *.
* J.B.S. Haldane. A defence of beanbag genetics. - "Perspectives in Biology and Medicine", 1964, v. 7, p. 343.
Генетика человека50-70-е годы XX в. отмечены значительными успехами в исследованиях генетики человека. Используя новые методы анализа, удалось обнаружить ранее неизвестные наследственные заболевания человека, определить природу их возникновения, разработать ряд методов изучения хромосомного комплекса и т.д. В наше время медицинская генетика использует в своих исследованиях пять основных методов: популяционно-статистический, клинико-генеалогический, близнецовый, цитогенетический и генетико-биохимический.
Анализируя историю исследований в области медицинской генетики, нельзя не отметить работы Холдейна по генетике человека, хотя в его время наука не располагала многими известными сейчас данными. Несмотря на то, что Холдейн не получил специального медицинского образования и иногда не в состоянии был проводить непосредственные исследования в области медицинской генетики, он очень хорошо знал все, что касалось наследственных заболеваний человека, отдав немало сил изучению этого вопроса. Во многих случаях им были сделаны принципиальные выводы, ускользнувшие от внимания других исследователей, специалистов в этой области; некоторые из них не утратили своего значения и по сей день. Так, Холдейн одним из первых применил математические методы для построения карты распределения генов в хромосомах человека. Он усовершенствовал методы точного изучения концентрации наследственных болезней в популяциях животных и также одним из первых приближенно определил процент мутирования нескольких генов у человека.
Холдейн придает большое значение влиянию факторов внешней среды на процесс формирования организма. В статье "Генетика и современные социальные теории" (1934), написанной по просьбе советского журнала "Успехи современной биологии", он отмечал:
"Люди отличаются друг от друга, и эти различия зависят частью от наследственных данных, частью от среды. Какой-либо ген одаренности может привести к совершенно различным результатам в зависимости от тех возможностей к образованию, которые имеются у соответствующего лица. Так, например, крестьянский сын, обладающий гениальным природным дарованием к математике, может не иметь случая проявить свои способности. И, наоборот, ребенок богатых родителей, который, будучи бедным, мог бы стать слабоумным, в результате индивидуального обучения может достичь среднего интеллектуального уровня. Поэтому генетик, желающий изучить главнейшие наследственные различия у животных или растений, должен содержать объекты изучения в однородной среде" *.Холдейн предупреждал о необходимости быть осмотрительными при экстраполяции на человека данных, полученных на животных, считая, что не следует спешить с практическими выводами: нужно методично и настойчиво накапливать фактический материал, обобщая его и подвергая тщательной теоретико-математической обработке.* Дж.Б.С. Холдейн. Генетика и современные социальные теории. - "Успехи современной биологии", 1934, №3, с. 426.Он осознал возможность перенесения ранее разработанных им математико-аналитических методов из области эволюции на популяционную генетику, в сферу изучения наследственности человека. Однако делал это он с большой осторожностью, что видно из его внимательного отношения к вопросу о генетическом равновесии, наблюдаемом в популяции человека при естественном отборе. Такой подход позволил ему еще в 1932 г. в работе "Факторы эволюции" показать возможность определения частоты возникновения мутаций, вызывающих высокую степень смертности у человека - носителя данного гена. На примере гемофилии, возникающей в результате наличия в X-хромосоме гена, определяющего развитие данного признака, Холдейн показал, что элиминация этого гена в каждом поколении из-за низкой жизнеспособности гемофиликов должна уравновешиваться новыми повторными мутациями, ибо в противном случае болезнь должна была бы вскоре исчезнуть. Он полагал, что законы естественного отбора не действительны в отношении нормальных женщин, гетерозиготных по данному гену, которые встречаются почти в два раза чаще, чем мужчины, обнаруживающие гемофилию.
Холдейн считал, что около трех случаев гемофилии на 100 тыс. человек возникает в результате мутации в каждом поколении. В случае появления гемофилии среди мужчин (один случай на 30 тыс.) темп мутаций этого гена в течение одного поколения составил бы около 10-3 или несколько выше.
Позднее, в 1953 г., Холдейн разработал общую формулу для генов, сцепленных с полом:
2m + v = (1 - f)x,
где m и v - темп мутаций у женщин и мужчин, х - частота гемофилии среди мужчин при рождении, f - выживающая часть мужчин, пораженных гемофилией, по отношению к общему числу больпых, принятому за единицу.
В 1938 г. в марксистском журнале "Modern Quaterly" был напечатай блестящий научно-популярный очерк Холдейна "Королевская кровь", включенный позднее в сборник статей для широкого читателя "Keeping Cool" ("Сохраняя хладнокровие"). В подзаголовке статьи значилось: "Исследование генеалогии гемофилии в королевских семействах Европы". В этой работе Холдейн критически использовал данные Р. Фишера о гемофилии и высказал свое несогласие с его мнением о том, что гемофилия появляется под влиянием инбридинга. Холдейн писал:
"Изучение большого количества материалов по наследованию гемофилии не подтверждает мнения Фишера... Иного рода дефекты физического и умственного порядка, которые прослеживаются в родословных некоторых королевских семейств, можно с большой долей вероятности приписать влиянию инбридинга, но вряд ли это относится к гемофилии".В том же 1938 г. американский журнал "Science and Society" опубликовал ответ Холдейна буржуазному экономисту А.П. Лернеру. Статья эта (не упоминаемая в библиографических обзорах работ Холдейна) интересна тем, что в ней Холдейн подчеркивает ту огромную роль, которую оказали на формирование его мировоззрения и методологию научного мышления произведения Энгельса, и относится она именно к периоду его интенсивных исследований в области популяционной генетики человека. Вот что он там пишет:"Должен признаться, что постепенное, постоянно углубляющееся изучение диалектического материализма воодушевило меня на осуществление большей части научных работ, опубликованных в последнее время. Например, недавно я опубликовал мое первое серьезное исследование о частоте мутирования генов человека. Я сделал это после того, как понял в процессе работы, выполняемой по просьбе Госиздата, над статьей «Современное состояние теории естественного отбора» * необходимость попытаться изложить менделевскую теорию эволюции в понятиях диалектики. За пять лет до этого я подверг математическому истолкованию вопрос о равновесии (или почти равновесном состоянии) между мутациями и отбором. Мне это стоило больших умственных усилий, в результате которых (спасибо Энгельсу!) мне удалось словесно выразить эту закономерность и убедиться в том, что это приближенное равновесие является ключом к пониманию многих удивительных фактов, в частности такого явления, как частота гемофилии в человеческих популяциях.Следуя традиции своего учителя Бэтсона, который полагал, что особое внимание следует в науке уделять исключениям ("Treasure your exceptions"), Холдейн заинтересовался двумя признаками, сцепленными с полом, - гемофилией и дальтонизмом, редко встречающимися в человеческих популяциях, и оценил частоту рекомбинаций между этими двумя генами.Я думаю, не случайно моя лаборатория является, вероятно, единственной в мире за пределами Советского Союза, где исследуется равновесие между мутациями и отбором. Изучение произведений Энгельса в течение нескольких лет способствовало кристаллизации в моем сознании нового принципа, основанного на объединении разрозненных данных, связанных с отбором у домашних животных.
Я не думаю, что без изучения трудов Энгельса я не пришел бы к таким результатам. Но мне хотелось бы подчеркнуть, что успехами моих исследований я во многом обязан мыслям Энгельса. И поскольку я убедился в том, что диалектический материализм является весьма действенным инструментом для научного творчества, считаю нужным заявить об этом..." **.
* Статья в русском переводе в печати не появилась.** Цит. по: Г.Э. Фельдман. Джон Холдейн о влиянии Энгельса на методологию научного мышления. - "Вопросы философии", 1970, № 11.

Дж. Холдейн (1940).
Первое измерение степени сцепления для случаев гемофилии и дальтонизма было осуществлено им в 1937 г. в совместной работе с Дж. Бэллом, когда уже было установлено, что гены, обусловливающие эти заболевания, находятся в Х-хромосоме. А еще в 1935 г. Холдейн, исходя из равновесия между мутациями и отбором, рассчитал и сформулировал концепцию о темпах мутации одного гена у человека. Иначе говоря, он установил то, что ныне принято называть косвенным методом определения темпа мутирования.
В 1947 г. Холдейн подсчитал, что темп мутирования гораздо выше у мужчин, чем у женщин, и что средняя его величина составляет 3,2 Х10-5 генов в поколении. Подобные наблюдения были проведены в отношении сцепленного с полом гена мышечной дистрофии *. Результаты определения темпа мутирования сцепленных с полом генов у человека с применением косвенного метода анализа на материале, собранном разными исследователями, оказались вполне сопоставимыми с темпом мутации, определенным при прямых наблюдениях над спорадическими случаями появления летальных признаков.
* J.B.S. Haldane. Mutation in the sex-linked recessive type of muscular dystrophy. A possible sex difference. - "Annals Human Genet.", 1956, v. 20, p. 344.Математически анализируя наследование резус-несовместимости, Холдейн показал *, что резус-фактор может находиться в состоянии нестойкого равновесия в тех случаях, когда частота встречаемости гена около 1/2, но что большинство обособленных популяций проявляет тенденцию превратиться в положительные или в отрицательные. Этот вид отбора, направленный на элиминацию гетерозигот, Холдейн рассматривал как явление, чуждое дарвиновскому принципу отбора. Он предположил, что отбор противоположного типа в пользу гетерозиготности, ведущий к стойкому равновесию, может способствовать объяснению широкого распространения некоторых генов, вызывающих болезни среди шшуляций человека. Холдейн считал, что эти болезни, как это имеет место при амавротической идиотии, микроцитемии, а также в случае серповидно-клеточной анемии, слишком часто встречаются, чтобы можно было их приписать давлению мутаций **. Впоследствии эти положения получили полное подтверждение во многих исследованиях.* J.B.S. Haldane. Selection against heterozygosis in man. - "Annals Eugen.", 1942, v. 11, p. 333.Холдейн поддерживал гипотезу, что хромосомные абберации у человека могут в значительной степени явиться причиной биологической неполноценности, в особенности у интерсексов *, причем было это им высказано задолго до цитологического подтверждения данного положения. Здесь же он предсказал возможность использования культур лейкоцитов человека для исследования его хромосом.** J.B.S. Haldane. The estimation of the frequencies of recessive conditions in man. - "Annals Eugen." 1938, v. 8, p. 255.
* J.B.S. Haldane. Genetical evidence for a cytological abnormality in man. - "J. of Genetics", 1932, v. 26, p. 341.Значительно позднее, в 1960 г., в небольшой статье "Группы крови и трисомия у человека" Холдейн обращается к генетикам и врачам Индии с призывом исследовать группу детей с болезнью Дауна и другие аномалии хромосом у человека. Как известно, трисомия встречается главным образом у детей, родившихся от матерей, возраст которых превышает 35 лет.В совместной работе с А. Рей "Генетика распространенной в Индии ненормальности пальцев" (1965), подготовленной к печати незадолго до кончины Холдейна, изложены результаты наблюдений над укороченным четвертым пальцем на ноге. Авторы показали, что этот признак вызывается аутосомным доминантным геном с неполным проявлением и отмечается в северо-восточных районах Индии.
Холдейн также установил вероятный новый доминантный признак, сцепленный с полом у человека *. Считалось, что дефект эмали, приводящий к потемнению зубов, наследуется во многих семьях как аутосомный доминантный признак. Однако Холдейн обнаружил родословные, где этот дефект вызывается, по-видимому, геном, расположенным в Х-хромосоме.
* J.B.S. Haldane. A probable new sex-linked dominant in man. - "J. Hered.", 1937, v. 28, p. 58.В исследовании "Равновесие при естественном отборе сцепленных с полом генов" (1964) Холдейн показал, что равновесие пары аллеломорфов в случае наследования сцепленного с полом гена может сохраняться при естественном отборе, если приспособленность мутантной гемизиготы и гомозиготы отличаются от нормального в противоположных направлениях, даже если женская гетерозигота менее приспособлена, чем гомозиготы в среднем. Но, если приспособленность мутантной гомозиготы и гемизиготы отличаются в одном и том же отрицательном направлении от нормальных, стабильность требует того, чтобы гетерозиготная самка обладала более высокой приспособленностью, чем нормальная форма.Холдейн много трудился над методами вычислений для обработки данных, полученных другими исследователями в области генетики человека. Его идея о корреляции между возрастом и проявлением изменений составила эру в изучении генетических модификаций *. Он предложил весьма оригинальный метод анализа возможных взаимодействий между наследственностью и средой **. Им также была показана корреляция между приспособленностью родителей и полноценностью потомства в условиях устойчивого равновесия с преимуществом гетерозигот ***.
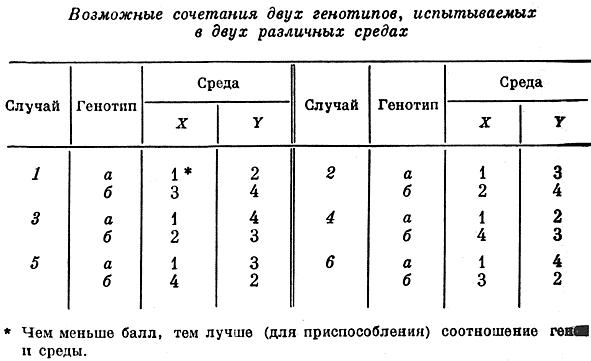
* J.B.S. Haldane. The partial sex-linkage of recessive spastic paraplegia. - "J. of Genetics", 1941, v. 41, p. 141.В работе "Взаимодействие наследственности и среды" (1946) Холдейн рассматривает различные климатические условия, в которых живет человек, и формы культуры в их историческом аспекте. Холдейн показал, что если людей разделить по их генотипу на две группы (а и б) и распределить по двум средам (X и Y), то возможны шесть резко различных реакций некоторых признаков, поддающихся количественному измерению и "градуированию" (вес, умственные способности, мышечная сила и т.п.).** J.B.S. Haldane. The interaction of nature and nurture. - "Annals Eugen.", 1946, v. 13, p. 197.
*** J.B.S. Haldane. Parental and fraternal correlations for fitness. - "Annals Eugen.", 1949, v. 14, p. 288.
В случаях 1 и 4 генотип а имеет преимущество по сравнению с генотипом б, однако если в случае 1 генотип б лучше проявляется в той же среде X, что и генотип а, то в случае 4 такого совпадения не наблюдается и б лучше проявляется в той среде, где а проявляется менее удовлетворительно.
В случаях 2 и 3 среда Y менее благоприятна, чем среда X для обоих генотипов; но в случае 2 в среде Y генотип а имеет преимущество перед б, а в случае 3 дело обстоит наоборот. Наконец, в случаях 4 и 6 наилучшим сочетанием оказывается генотип а и среда X; генотип б лучше проявляется в среде Y; имеются различия в реакции генотипов при остальных двух сочетаниях (см. таблицу). Холдейн подчеркивает, что для большинства этих случаев можно было бы привести примеры и из агрономии и животноводства.
Он доказывает, что если взять три генотипа и три среды, то число возможных реакций составит 10080. При этом он дает общую формулу для подсчета реакций:
(mn) / m!n! ,
где m - число различных генотипов, n - число различных сред. При увеличении m и n число типов реакций возрастает с удивительной быстротой; так, например, при m = n = 10 число типов реакций равно 7.09 X10144. Исходя далее из того, что поведение различных генотипов в различных средах показательно в отношении не одного признака, а их совокупности (например, если одновременно учитывать умственные способности, продолжительность жизни и иммунитет к заболеваниям), Холдейн подсчитал, что даже в простейшем случае, когда имеется два генотипа, две среды и два признака, можно наблюдать 72 различных типа взаимодействия. При этом он дает общее выражение для различных комбинаций взаимодействия:
(mn)! (k/m)! m! n! k! , где k - число различных критериев.
Следует иметь в виду и такое важное обстоятельство, что генетически различные группы могут в разных средах дать одинаковые результаты. По мере усложнения компонентов и видов взаимодействия число различных типов реакций сильно возрастает. Холдейн приходит к выводу о том, что какие-либо обобщения относительно "превосходства" одной группы людей над другой или одного человека над другим следует делать очень осторожно.
В исследовании "Новый тест достоверности при получении данных из ограниченных популяций в приложении к инбридингу у человека" (1963) Холдейн и Джакар дают математическое выражение распределения средних вариантов и ковариантов и корреляции в опыте в условиях ограниченной популяции. Они полагают, что их метод более точен, чем метод Стюдента для определения значения разности между средними величинами.
Последние годы своей жизни в Индии Холдейн руководил исследованиями по генетике человеческих популяций, в частности в штате Андхра-Прадеш, где коэффициент инбридинга среди прибрежных популяций, насчитывающих свыше 10 млн. жителей, превышает 0,02. Он считал, что развивающиеся страны должны предпринимать доступные им исследования в тесном сотрудничестве с квалифицированным научным персоналом передовых стран. Холдейн планировал работы по сцепленной с полом цветной слепоте, изучению наследования сцепленного с Y-хромосомой гипертрихоза ушей, глухонемоты и склонности к леворукости, а также генетические исследования местных популяций, например, изучение частоты дефекта активности глюкозо-6-фосфатдегидрогеназы в малярийных районах штата Андхра-Прадеш. Но смерть помешала Холдейну осуществить эти планы.
Таким образом, Холдейн был первым ученым, заложившим основы демографической генетики как самостоятельной дисциплины. Будучи в основном ученым-теоретиком, сам он не принимал непосредственного участия в изучении человеческих популяций, и тем не менее его идеи оказали весьма плодотворное воздействие на успехи демографической генетики. Его мысль о синтетическом объединении достижений из области генетики человека с математико-статистическими моделями, разработанными применительно к демографии, открывает новые пути для использования количественного подхода к науке о человеке.
Идеи, имеющие непосредственное отношение к демографической генетике, были изложены Холдейном в 10 сообщениях по математической теории эволюции, которые печатались в период с 1924 по 1934 г. В этих работах был сформулирован основной круг вопросов, подлежащих изучению и определены конкретные случаи отбора, которые требуют дальнейшего исследования; наконец, сделана первая попытка применить законы менделевской генетики к демографическому материалу, чтобы объединить их в единую науку.
На основании тщательного математического анализа Холдейн пришел к следующему выводу: оценка замедленного или ускоренного темпа отбора и его эффективности во времени в пределах обособленной популяции может быть успешно осуществлена лишь после того, как исследователь будет располагать материалами об особенностях наследования данного признака, об особенностях и системах размножения конкретных организмов, о наличии отбора (касающегося представителей того или иного пола или у того и другого), об интенсивности отбора и, наконец, о темпе прогрессивного развития или угасания отдельного признака в отдельных группах организмов.
Холдейн неоднократно указывал на то, что эффективные и действенные исследования в науке о народонаселении возможны лишь на стыке ряда дисциплин: философии, социологии, социальной гигиены, экономической географии, истории, демографической статистики, социальной психологии, этнографии и физической антропологии и таких разделов биологии, как генетика и физиология. Он часто возвращался к классическим трудам по естественному отбору у человека, например к работам К. Пирсона о наследовании долголетия, Р. Фишера о наследовании плодовитости, М.Н. Карна и Л.С. Пенроуза об отборе на вес при рождении, и беспощадно критиковал * работы, в которых замалчивались эти классические исследования.
* См., например, его статью "Parade of ignorance" ("Парад невежества") - "J. of Genetics", 1962, v. 58, p. 138.На основе данных Пенроуза о весе младенцев при рождении и о смертности среди новорожденных в Лондоне Холдейн вычислил интенсивность естественного отбора, которая оказалась равной приблизительно 2,4%, и высказал гипотезу о повышенной плодовитости тех индивидов, вес которых при рождении превышал на 1 кг оптимальную величину, рассматривая это как компенсацию за повышенную смертность среди таких крупных новорожденных (1961).Холдейн видел перед современной медицинской генетикой следующие задачи:
1. Дифференцировать врожденные заболевания, обнаруженные при рождении или вскоре после него. Например, иногда имеет место врожденный сифилис, который развивается вследствие пренатального заражения. И есть группа заболеваний, которая в значительной степени также зависит от ненормальных условий пренатального периода, но где причастен и элемент наследственности (сужение привратника, монголизм и др.), что поддается контролю лечебно-профилактическими мерами в пренатальный период, например введением соответствующих гормонов в материнский организм.Джон Бернал в письме к автору настоящей книги от 8 мая 1967 г. писал:2. Различать внешне сходные заболевания, вызванные различными факторами, поскольку даже различные формы одной болезни требуют различных профилактических и терапевтических мероприятий.
3. Отличать факторы генетического порядка от факторов внешней среды при изучении наследования таких, например, заболеваний, как рахит или туберкулез.
"Эволюцию научных интересов Холдейна следует рассматривать в развитии от физиологии к биохимии с выходом в генетику. На этом пути он принял активное участие в дискуссии по вопросу о взаимодействии наследственности и среды, придавая большое значение роли внешнего социального фактора. Он никогда не был сторонником и последователем школы менделевского детерминизма и резко выступал против применения учения Менделя к человеческим популяциям. С самого начала научной деятельности Холдейн выступал против мистических представлений и боролся с ними".При публикации научных работ, исходящих из его лаборатории, Холдейн неизменно отдавал должное (а часто переоценивал) роль того или иного из своих сотрудников в постановке отдельных исследований. К сожалению, оставшиеся после него работы содержат лишь часть тех бесценных идей, которыми он щедро делился со своими учениками и коллегами.Обобщив и подвергнув математической обработке данные, собранные многочисленными исследователями, Холдейн обогатил теорию новыми генетическими идеями, используя для этой цели тончайшие аналитическое методы. Профессор С.А.Б. Смит, ныне возглавляющий кафедру биометрии Лондонского университета, отмечает, что
"Холдейн обладал удивительным даром проникать в самую сущность генетических явлений. Он не страдал «болезнью предвзятых идей» и от его взора не ускользало многое из того, мимо чего проходили его коллеги по профессии. Он всегда с готовностью прислушивался к критическим замечаниям о том, что он в тех или иных своих построениях не был прав. Если кто-нибудь обнаруживал ошибку в его работах, он, как правило, говорил: «Вы правы, мы опубликуем это в виде совместного сообщения». Он никогда не воспринимал критику по вопросам науки как личный выпад в его адрес и, если он публично выступал с критикой своих коллег, то всегда строго придерживался фактической стороны дела" *.Выступая в 1963 г. на XI Международном конгрессе генетиков в Гааге с докладом "Значение современной генетики для человеческого общества", Холдейн говорил: "Необходимо, чтобы широкие массы людей знали основы генетики человека, поскольку этот вопрос касается всего человечества. Незнание может дать простор для пропаганды предрассудков или псевдонаучных теорий. Задача ученых - разъяснять и пропагандировать основу знания и неустанно развивать науку о наследственности".* Cedric А.В. Smith. "Ass. of Scientific Workers Journal", 1965, v. 11, №2.Радиационная генетика
Прогноз генетических повреждений, индуцируемых радиацией, и разработка биологических мер защиты от вредного действия радиации являются одной из наиболее актуальных проблем современной радиационной генетики.
Работы Холдейна в области радиационной генетики появились за шесть лет до взрыва атомной бомбы в Нагасаки и Хиросиме. 18 мая 1939 г. в газете "Дейли Уоркер" была напечатана сенсационная для того времени статья - "Перспективы использования атомной энергии". В ней Холдейн на основании тщательного анализа новейших успехов ядерной физики предсказывал реальную возможность создания в ближайшие годы атомной бомбы и рассматривал почти все аспекты, связанные с разработкой атомной энергии.
В декабре 1964 г., после кончины Холдейна, эта статья была вновь напечатана в "Дейли Уоркер", чтобы показать удивительный дар его научного предвидения. Фактический материал, позволивший Холдейну сделать столь ответственные обобщения, был опубликован в журнале "Nature" в серии сообщений физиков-теоретиков в конце 1938 - начале 1939 г.
Холдейн не ограничился, однако, общими научными прогнозами, как генетик он попытался решить некоторые вопросы, связанные с воздействием ионизирующего излучения на наследственный аппарат живых организмов.
В работе 1947 г. "Вредный эффект индуцированных рецессивных мутаций" Холдейн, используя исследования Г. Мёллера *, попытался показать, что видимые эффекты от рецессивных мутаций, обусловленных радиацией, достигнут своего максимума спустя четыре-пять поколений и будут сохраняться приблизительно на одном н том же уровне в течение достаточно долгого времени. В будущем каждая вредная рецессивная мутация будет обусловливать гибель определенного количества зигот. Эта элиминация может происходить различными путями. Если ген летальный, то гибель произойдет на очень ранней сгадии развития и, возможно, приведет к понижению плодовитости. На более поздних стадиях вредные мутации могут обусловить аборты, мертворождение и смертность в раннем возрасте. И, наконец, подобная мутация может повести к понижению жизнеспособности организма, например человека, в среднем на 5%. Холдейн считает, что если данная популяция окажется облученной, то продолжительность жизни уменьшится более чем наполовину. Это будет относиться ко всем мутациям, которые в основном понижают жизнеспособность гомозигот и тем самым элиминируются из популяции.
* Н.J. Muller. The role played by radiative mutation in mankind. - "Science", 1941, v. 93, p. 438.Используя ряд математических расчетов, Холдейн приводит данные по возникновению мутации в человеческой популяции в результате взрыва атомной бомбы при условии, что каждый индивидуум получит в среднем дозу в 20 рентген. Общее количество возникающих рецессивных летальных мутаций с учетом данных, полученных на дрозофиле, будет около 4000. Холдейн, однако, высказывает предположение, что это количество должно быть увеличено по отношению к человеку, поскольку он имеет большее число генов, чем дрозофила, а также в силу его более высокой радиочувствительности. Возникшие мутации будут обусловливать гибель людей в течение многих поколений, и, по-видимому, на каждое поколение около 20 смертей из 100 000 будут результатом облучения.Для оценки генетического действия радиации на человека необходимо располагать рядом данных: о величине дозы, удваивающей спонтанную частоту мутаций, об относительной генетической радиочувствительности как половых, так и соматических клеток, о роли фактора времени в генетическом эффекте действия радиации (т.е. было ли облучение острым или хроническим, однократным или фракционированным). Все эти данные необходимо получить в отношении разных видов ионизирующих излучений и по разным типам мутаций (генные, хромосомные). Но и в настоящее время эти вопросы в отношении человека еще не полностью изучены.
В связи с этим представляет, безусловно, интерес крунианская лекция Холдейна "Формальная генетика человека", прочитанная им в 1948 г. в Лондонском королевском обществе, где он впервые выдвинул ряд положений, на основании которых можно высчитать минимальную удваивающую дозу. Холдейн исходил из аналогии между закономерностями у человека и у других организмов, в частности у дрозофилы и мыши. Сущность его подхода состояла в оценке доли частоты спонтанных мутаций у человека, которую можно, вероятно, приписать действию излучения естественных источников. Он полагал, что если принять: f - часть спонтанных мутаций, обусловленная действием излучения естественных источников, r - мощность дозы, создаваемой естественным фоном (r в день), m - частота мутаций, вызываемых облучением дозой в 1 r,) s - частота спонтанных мутаций, t - средний возраст особей, участвующих в размножении (дни), для дрозофилы, то
f = (mrt) / s
Для случая человека те же величины Холдейн обозначил соответствующими прописными буквами:
F = (MRT) / S
К этой формуле нередко прибегают, причем следует отметить, что частоты спонтанных и индуцированных мутаций (s и m) у дрозофилы установлены более точно, чем у человека (S и М).
В 1955 г. Холдейн вновь возвращается к этому вопросу *. В том же году он выступает с полемической статьей "Генетический эффект атомных взрывов" против Дж. Кокрофта, полагая, что тот недооценивает опасности генетического эффекта радиации **.
* J.B.S. Haldane. Genetical effects of radiation from products of nuclear explosions. - "Nature", 1955, v. 176, p. 115.Холдейн считал, что доза, удваивающая частоту мутаций у человека, несколько превышает 3 рентгена на поколение, а не 50 рентген, как полагал Джон Кокрофт.** J.B.S. Haldane. The genetic effects of atomic bomb explosions. - "Current science", 1955, v. 24, p. 399.
"Я умышленно, - пишет Холдейн, - использовал расчеты, полученные до того, как возник вопрос о генетическом эффекте, вызванном экспериментальными взрывами. Более поздние расчеты могли быть продиктованы сознательными или неосознанными политическими предубеждениями. Исследование этого вопроса требует исключительно осторожного планирования с учетом того обстоятельства, что многообещающие программы исследований могут оказаться излишними. До опубликования результатов такого рода исследований многим биологам трудно будет разделить оптимизм сэра Джона Кокрофта".Далее Холдейн указывает на то, что Коккрофт не учитывает генетическую опасность радиоактивных осадков в таких слаборазвитых странах, как Индия, Пакистан, Индонезия, в странах тропических районов Африки, Бразилии, где население большую часть времени проводит на воздухе, а их жилища отличаются тонкими стенами и не защищены от гамма-облучения."Я уверен, - пишет он, - что Кокрофт сожалеет об этом, но некоторые, связанные с ним лица, могут и не разделять его чувств. Так, например, сэр Эрнст Рок Карлинг, официальный представитель Британского правительства на недавно состоявшейся Женевской конференции заявил:На Международном генетическом симпозиуме в Японии в 1956 г. Холдейн предложил следующие этапы исследований для более точного определения частоты мутаций, возникающих под влиянием облучения у мышей.«С точки зрения планирования население земного шара может превысить в будущем количество потребляемых пищевых продуктов и в этом случае, может быть, не следует уж очень сожалеть о снижении плодовитости и сокращении длительности жизни человека. Еще меньше хлопот в отношении стран тропического пояса».
Интересно отметить, что Научный комитет ООН по действию атомной радиации на человека в результате детального рассмотрения и обсуждения данных по генетическому действию радиации пришел к выводу, что доза, удваивающая спонтанный темп мутации у человека, лежит между 10-100 рентгенами. Очевидно, что данные оценки величины мутагенного эффекта радиации приблизительны, и потребуются еще многие и многие исследования по радиационной генетике млекопитающих, чтобы получить более точную оценку генетического действия радиации на человека".
1. Изучение влияния облучения на соотношение полов. Сопоставление данных в отношении мышей с данными в отношении человека, собранными в разных странах, для сравнения мутабильности этих двух видов.Понятие "генетического груза" позволило Холдейну в 1957 г. подойти к оценке суммарного влияния мутаций на популяцию, что приобретает все большее значение для изучения влияния радиации и мутагенных химических веществ на частоту возникновения мутаций у человека.2. Изучение видимых мутаций, включая летальные, действующих на поздней стадии развития зародыша или после рождения.
3. Изучение возникновения транслокаций и инверсий с помощью анализа их влияния на плодовитость и частоту нарушения сцепления (эти изменения могут наблюдаться при воздействии большими дозами и в короткие сроки).
4. Установление рецессивных леталей, сцепленных с полом, путем использования сцепленных с полом маркеров.
5. Изучение частоты мутации отдельных генов.
6. Учет аутосомных рецессивных леталей с помощью метода Холдейна, разработанного им для анализа возникновения рецессивных леталей (1956).
7. Обнаружение мутаций методом Грюнберга, разработанным им для анализа повышения частоты мутаций признаков скелета у мышей (1954).
8. Использование статистических методов обработки данных о длительности жизни, плодовитости, темпе роста и т.д. у потомства мышей, подвергнутых облучению, что позволит выявить мутации, повышающие жизнеспособность, поскольку естественный отбор элиминирует гены, снижающие жизнеспособность.
В работе 1956 г. "Определение темпа мутаций под влиянием ионизирующей радиации у млекопитающих" Холдейн выступает как воинствующий гуманист:
"... это сообщение опубликовано в Индии, - пишет он, - надеюсь, оно полностью будет воспроизведено и в других странах по следующей причине. Описанный метод может быть успешно использован одной из держав, производящих атомные бомбы, или в стране, связанной с такой державой. В этом случае неизбежно проявится тенденция либо к запрещению опубликования подобных результатов исследования, что раскрыло бы опасность атомных бомб для населения всего мира, либо стремление преуменьшить значение полученных данных. Надо думать, что в Индии или Японии проявится стремление к преувеличению значения этих данных. Если результаты, полученные в странах того и другого типа, в основном совпадут, то это и будет отражением истинного положения вещей".В 1958 г. Холдейн еще раз возвращается к этой теме в лекции "Генетический эффект квантов и частиц высокой энергии", прочитанной в Калькутте.В ходе экспериментальных работ по изучению интимных механизмов, связанных с регуляцией щелочности и кислотности внутренней среды человека, у молодого Холдейна возникла потребность глубже вникнуть в сущность жизненных явлений и проследить за ходом тончайших процессов, происходящих в целом организме. Эта потребность диктовалась также и его интересами, лежащими в области генетики.
В 1920 г. он изложил теорию "один ген - один фермент" *, возникшую под влиянием идей, шысказанных еще в начале века Гарродом (1902) ** и Л. Кэно (1903) ***, а к 1922 г. Холдейн был уже автором нескольких математических исследований по генетике. Молодого ученого увлекает разгадка биохимической природы гена. Он решает срочно овладеть основами биохимии и не штудированием учебников и прослушиванием лекций, а участием в экспериментальной и практической работе; пожертвовав каникулярным временем, летом 1921 г. он поступает в лабораторию клинической биохимии при Эдинбургском госпитале. Со студентами-медиками он участвует в обходе больных, но большую часть времени проводит в лаборатории, осваивая технику экспериментов, при этом занятия его тесно связаны с теми разделами физиологической химии, которые непосредственно или отдаленно касаются сущности механизмов наследственности и химии обменных процессов ****.
* J.B.S. Haldane. Some recent work on heredity. - "Trans. Oxford. Univ. Jr. Sci. Club", 1920, series 3, v. 1, p. 3.Исследования Холдейна, посвященные изучению природы развития ацидоза и алкалоза, опубликованные в 1920-1923 гг. в физиологических журналах, а также его первые работы по сцеплению признаков заинтересовали Ф.Г. Гопкинса (1861 -1947), одного из крупнейших биохимиков того времени. После открытия триптофана Гопкинс интенсивно работал над изучением роли окислительно-восстановительных и энергетических процессов в мышечном сокращении и в 1921 г. возглавил лабораторию биохимии в Кембриджском университете. Его исследования путей образования молочной кислоты и обнаружение ее ничтожных количеств в покоящейся мышце (в период сокращения) в присутствии кислорода опровергли господствующую в то время теорию депонирования внутримышечного кислорода. Заслугой Гопкинса было и то, что он впервые показал невозможность поддержания жизни животных при наличии в пище лишь белков, жиров и углеводов. С его именем связано создание учения о витаминах - вспомогательных пищевых факторах, как он их назвал, без которых невозможна нормальная жизнедеятельность.** A.E. Garrod. Inborn errors of metabolism. Oxford, Univ. Press, 1902.
*** L. Cuenot. Hypothese sur l'heredite des couleurs dans les croisements des souris noires, grises et blanches. - "C. R. Soc. Biol.",
1903, v. 55, p. 301.**** Возможно, Бэтсон имел в виду именно Джона Холдейна, когда однажды с грустью заметил: "Все одаренные молодые ученые изменяют генетике в пользу биохимии".

Фредерик Гопкинс
Для понимания эволюции интересов Хблдейна особенно важен тот факт, что именно в этот период Гопкинс задался целью изучить врожденные нарушения обмена веществ у человека. В 1922 г. он предложил Холдейну занять в его лаборатории место лектора по биохимии. С 1923 г. Холдейн приступил к чтению курса общей биохимии и раздела энзимологии, став вторым человеком в лаборатории после Гопкинса, и проработал с ним бок о бок десять лет - до 1932 г. Вспоминая об этом периоде, Холдейн писал: "Основной смысл моей деятельности состоял в том, что я руководил научной работой ряда сотрудников, работающих в самых разнообразных областях, включая физиологическую химию человеческого организма, животных, высших растений и бактерий".
Личные интересы Холдейна в области биохимии в этот период (1923-1930 гг.) сосредоточились на ферментах - основных катализаторах, координирующих развитие обменных процессов. Хотя к этому времени роль ферментов в метаболизме была установленным фактом, однако предстояло изучить специфику и диапазон их действия, функции и пути их реализации, а также создать научно обоснованную систему их классификации. Эти вопросы приковали к себе внимание Холдейна на долгие годы. В 1925 г. в совместной работе с Дж.Е. Бриггсом Холдейну удалось показать, что кинетические реакции ферментов и степень сродства их к субстрату подчиняются известным законам термодинамики. При помощи математического аппарата Холдейн разработал уравнение скорости развития некоторых ферментативных реакций.
В своих исследованиях Холдейн постоянно стремился соотнести локальные биохимические процессы с событиями, разыгрывающимися в целом организме, т.е. работая в области биохимии, он оставался физиологом. Термином "механизм" он пользовался с оговорками и скептически относился к многочисленным попыткам своих коллег переносить данные, полученные в динамических опытах in vitro, к явлениям in vivo.
В 1930 г. он опубликовал монографию "Энзимы" - сильно расширенный и переработанный для печати цикл лекций, прочитанных им в Кембридже в 20-х годах. Холдейн считал, что эта книга в значительной степени является дальнейшим развитием положений В. Бейлиса ("Природа действия энзимов", 1923), в которой тот отстаивал взгляд на энзимы как на катализаторы. В 1931 г. по инициативе крупного биохимика и генетика Курта Штерна книга была переведена на немецкий язык, а в 1934 г. - на русский (редакция и перевод С.И. Пронина).
Холдейн предполагал, что его читатель обладает значительными знаниями органической и физической химии, а также химии углеводов и высшей математики. Поэтому книга отличается лаконичностью, а математическая сторона представляет определенную трудность для понимания, поскольку автор, как правило, опускает довольно сложные промежуточные преобразования и дает сразу конечный результат.
Оценивая достоинства монографии "Энзимы", Л. Михаэлис в предисловии к немецкому изданию отмечал, что по насыщенности мыслями и фактическим материалом она стоит в одном ряду с такими солидными руководствами, как "Ферменты и их действие" К. Оппенгеймера и "Химия энзимов" Эйлера.
Книга Холдейна появилась в начальный период исследований в области энзимологии, в момент, предшествовавший кинетической стадии, когда стадия химических исследований еще не наступила, поскольку не была ясна природа ферментов. Холдейн на основе вполне допустимых предположений вычислил чистоту ферментных препаратов, приготовленных производственным способом, время жизни ферментно-субстратных комплексов и скорость реакций.
С ходом развития энзимологии взгляды Холдейна в некотором отношении, естественно, устарели. Но заслуга его, состоящая в том, что он придал этому разделу биохимии конкретность, осталась. Характерно, что Холдейн предвидел значение действия специфических ферментов в аналитических и химических процессах и высказал предположение, что изучение кинетики действия ферментов может способствовать пониманию их структуры, что было для того времени поистине пророчеством. Теперь мы знаем, что по концентрационным кривым активности ферментов можно судить, построены они из одной или нескольких субъединиц.
Глубокое знание всего, что было тогда известно о ферментах, и широкая осведомленность в вопросах генетики позволили Холдейну синтезировать обе эти важнейшие биологические науки.
Индийский биохимик профессор Д.П. Бурма в статье, посвященной памяти Холдейна, писал:
"Всего лишь два-три десятилетия назад в науке стали упоминать о генах и ферментах. Трудно представить себе кого-нибудь, кроме Холдейна, кто как он не только имел возможность поставить на ноги эти молодые науки, когда они еще находились в эмбриональном периоде, но и наблюдать, как они сейчас созрели. Интимное родство между генами и ферментами - установленный факт. На стыке биохимии и генетики родилась молекулярная биология. Холдейн относился к тем избранным, которые могли свободно вторгаться из одной области естествознания в другую и зорко следить за развитием и частного, и целого, что доступно лишь немногим" *.Весьма важно, что физико-химическая трактовка изучаемых биохимических явлений дана Холдейном в полном соответствии с успехами физической химии в тот период. Однако несмотря на интенсивное развитие физической химии за последние сорок лет, в первую очередь в области кинетики и катализа, изучения структуры веществ биохимического происхождения и исследования механизма взаимодействия веществ с излучением, книга Холдейна не утратила своего дидактического значения и с соответствующими изменениями и дополнениями для специалиста, работающего в области динамической энзимологии, может представить интерес и в наше время. Современные представления о теории активации и об основных чертах каталитических процессов своими корнями связаны с этой работой Холдейна.* "Science reporter" (New Dehli) Haldane number, 1965, p. 514.В предисловии, написанном в мае 1964 г., к американскому переизданию книги "Энзимы" Холдейн так характеризует современное значение книги.
"Переиздание книги преследует четыре цели:
во-первых, позволяет заинтересованным специалистам ознакомиться с уровнем знаний в своей области в недавнем прошлом. Работникам в энзимологии не безынтересно знать, что было и чего не было установлено в науке к 1930 г.;Думается, что с момента выхода моей книги энзимология развивалась в трех основных направлениях:во-вторых, может способствовать выяснению, какие направления исследований не получили должной систематической разработки и нуждаются в дальнейшем плодотворном развитии. Таблицы II (оптимальные значения рН для многих энзиматических реакций), V (концентрации субстрата, дающие половину максимальной скорости) и XII (температурные коэффициенты некоторых энзиматических действий), казалось бы, невольно ведут к обобщениям, которые, быть может, окажутся ложными, а с другой стороны, возможно позволят по-новому осветить общие вопросы, связанные с действием ферментов. Если бы я ушел на покой и перестал бы работать в науке, а этого еще не случилось, и к тому же жил бы в глуши, где к моим услугам была бы отличная библиотека, я бы попытался расширить содержание этих таблиц до объема отдельной монографии;
в-третьих, книга может оказаться полезным источником по библиографии, справочным аппаратом, по исследованиям прошлых, ныне нацело забытых лет; может при этом статься, что некоторые работы того времени будут иметь отношение к науке сегодняшнего дня;
в-четвертых, она может быть воспринята и как Серьезное Предупреждение. Читатели, возможно, воскликнут: «Как мог Холдейн, у которого изложение материала по кинетике ферментов в главах III и V требует многих дополнений (но лишь незначительных поправок!), ставить не на ту лошадь, как он делает это в некоторых последних главах книги». Возможно, что некоторые нынешние специалисты, молекулярные биологи, могут оказаться столь же близорукими, как я, и сейчас, 34 года спустя.
во-первых, очистка многих ферментов. Начиная с первой работы Дж.Б.Семнера установлено, что все ферменты являются белками и что многие лишены простетической группы;В ту пору я глубоко верил в то, что 30 лет спустя мы будем знать точную структуру активных центров ряда ферментов и исходя из этой структуры сможем составить ясное представление о свойствах ферментов. Моя мечта еще не сбылась. Частично это можно объяснить тем, что активные центры, видимо, включают в себя аминокислотные остатки ряда близлежащих и слабо между собой связанных пептидных цепей.во-вторых, в некоторых случаях мы знаем, а в ряде случаев можем с большой долей вероятности предположить, что структура фермента точно определяется структурой одного или более генов;
и, наконец, в-третьих, нам известно, что метаболические процессы - такие, как окисление и синтез, представляют собой серии катализируемых ферментом реакций. Реакции эти обратимы, поскольку они вызывают незначительное изменение свободной энергии. В них и вовлекаются малые молекулы, часто нуклеотиды, подобные аденозинфосфатам и фосфопиридиновым нуклеотидам, которые в 1930 г. еще не были известны.
Мечтаю дожить до того времени, когда смогу прочесть работу, в которой автор, исходя из последовательности аминокислот в двух ферментах объяснит, почему один из ферментов должен быть эстеразой, а другой - пептидазой...
... Такое переиздание лишний раз убеждает мепя в том, что я не зря прожил жизнь" *.
* J.B.S. Haldane. Enzymes. Massachusetts Institute of Technology, M. I. T. Press, Cambridge, Mass., 1964.Профессор Рене Вюрмсер (которого Холдейн в предисловии к "Энзимам" благодарит за ценные советы при подготовке книги к печати), говорил, что на этой книге воспитывалось, по крайней мере, два поколения современных биохимиков. После смерти Холдейна он вспоминал о нем:"Мы всегда в пем видели друга французской науки. Он часто приезжал с лекциями в Сорбонну, в Институт физико-химической биологии, в Институт Анри Пуанкаре. Он обладал редким даром приходить в восторг от научных успехов и открытий своих коллег и страстно писал и говорил об этих работах, дополняя и развивая их за счет своего удивительного воображения.До конца жизни Холдейн сохранял чувство благодарности к своим учителям. В предисловии, написанном в мае 1964 г. к американскому переизданию книги "Энзимы", он писал:Еще в ту, теперь далекую, пору конца 1920-х годов, Холдейн умел постигать тайны природы, доводя их до молекулярного уровня. За формальным выводом математических формул всегда проглядывал азартный поиск глубоко запрятанных причинных механизмов, направляющих течение процесса" *.
* "Haldane and modern biology". Baltimore, J. Hopkins Press, 1968, p. 315."Сэр Фредерик Гопкинс был человеком необыкновенной скромности и главным редактором той научно-монографической серии, в которой эта книга впервые увидела свет. Поэтому я не мог в предисловии 1930 г. выразить, как многим я ему обязан. До того как я начал работать в его лаборатории, я пользовался ферментами лишь в качестве аналитического инструмента и не придавал им никакого значения. Гопкинс убедил меня в том, что ферменты представляют собой центральную проблему биохимии. А мой отец, Джон С. Холдейн, примерно в 1910 г. установил, что, хотя гемоглобин не что иное, как гигантская молекула, его реакции можно предсказать на основании законов, относящихся к малым молекулам. Мне же оставалось лишь свести воедино концепции этих двух выдающихся ученых и разработать теорию действия ферментов, которая в основных своих положениях, хотя и в несколько схематической форме, подтвердилась".
Теория о происхождении жизни на ЗемлеВ конце 1928 г. *, а затем в 1929 г. ** Холдейн изложил свою гипотезу о вероятном механизме процесса происхождения жизни на Земле в лаконичном очерке "Возникновение жизни". Выступление Холдейна с обоснованием своей теории происхождения жизни явилось вполне закономерным, поскольку к этому времени он проявил себя как экспериментатор и теоретик почти во всех областях биологических наук (генетика, математическое обоснование эволюционного процесса, энзимология, космическая биология), без чего немыслима попытка подойти к решению проблемы происхождения жизни на Земле. Обращение к этой проблеме было также продиктовано глубоким интересом ученого к философским проблемам, к поиску теоретических решений генезиса живой материи с использованием данных из различных областей естествознания. В этот, короткий по форме, но блестящий по содержанию очерк Холдейн сумел вместить все основные концепции, связанные с проблемой происхождения жизни.
* Anonymous. News and views. - "Nature", 1928, v. 122, p. 933.К проблеме предбиологической эволюции он подошел с позиций Дарвина. Эта проблема обосновывалась и последовательно разрабатывалась им и в дальнейших работах. Весь ход рассуждений Холдейна построен на признании постоянного изменения как среды обитания организмов, так и их собственной внутренней среды, что полностью отрицает существование каких-либо сверхъестественных сил. "В прошлом примитивный организм мог возникнуть только из другого, сходного организма; или если предположить здесь действие какого-то естественного или сверхъестественного агента, то он должен был быть, по крайней мере, столь же разумным, как мы сами, и притом гораздо более знающим" *, - пишет он.** Дж.Б.С. Холдейн. Возникновение жизни. - В кн.: Дж. Бернал. Возникновение жизни. М., "Мир", 1969, с. 295-303.
* Цит. по кн.: "Происхождение предбиологических систем". М., "Мир", 1966, с. 20.По мнению английского биохимика Н. Пири, со времен Т. Гексли (1868) и Дж. Тиндаля (1874), когда первый выступил в защиту физической основы жизни, а второй решительно отверг религиозную точку зрения, заявив, что возникновение и развитие жизни - это естественный процесс и для его объяснения не требуется привлечения сверхъестественного творческого начала, еще не было такой четкости и ясности мысли при формулировке основных проблем, стоящих перед наукой о происхождении жизни, какую обнаружил Холдейн. Его теория о возможном механизме возникновения предбиологических систем из органических соединений покоилась на обширных знаниях из области эволюционной биохимии - науки, которая в ту пору еще не оформилась в самостоятельную дисциплину.Сообщение Холдейна было встречено с интересом, по крайней мере биохимиками; новизна предложенной гипотезы получила такое дружное одобрение, что Холдейн был несколько смущен и в работах (50-60-х годов) шутливо заметил, что сам начинает сомневаться в приемлемости своей гипотезы, поскольку он всегда с недоверием относился к ортодоксальности.
Ниже мы излагаем основные положения теории Холдейна, которую он развивал и дополнял в течение всей своей жизни по мере того, как периодически возвращался к ней. На протяжении 35 лет каждая из его статей по вопросу о происхождении жизни содержала, по крайней мере, одну новую гипотезу.
Классификация теорий происхождения жизни. В малоизвестной работе 1954 г. "Пути возникновения жизни" Холдейн так классифицирует теории происхождения жизни на Земле.
I. Материя и жизнь всегда существовали на нашей планете. Никакого начала не было (теория вечности жизни).Холдейн допускает правомочность и вероятность первой гипотезы. Однако считает, что если третья гипотеза окажется лишенной оснований, а четвертая - маловероятной, то только в этом случае первая гипотеза потребует самого пристального изучения.II. Жизнь - продукт деятельности сверхъестественного начала (витализм).
III. Жизнь развивалась постепенно, эволюционно, в результате медленных химических реакций.
IV. Возникновение жизни есть результат "невероятного события", неизбежность которого была почти предопределена во времени, при условии, что предварительно образовалось абиогенным путем достаточное количество материи соответствующего химического состава и соответствующего состояния. Слово "почти" имеет строго математический смысл. В данном контексте оно означает, что, каким бы малым положительным числом ни пользоваться, всегда можно определить отрезок времени, прежде чем вероятность неслучившегося события станет меньше названного числа.
Дж. Бернал основную ценность представлений Холдейна усматривал в том, что им были "впервые сведены вместе две главные идеи. Это, во-первых, представление о существовании промежуточных форм, которыми Холдейн считает вирусы - и, в частности, бактериофаг Д'Эрреля, и, во-вторых, идея об атмосфере, первоначально не содержащей кислорода" *.
* Дж. Бернал. Возникновение жизни, с. 304.Условия первичной Земли. По мнению Холдейна *, жизнь на Земле в своем современном виде могла возникнуть лишь тогда, когда во Вселенной возникла адекватная химическая обстановка, при которой кометы обогащали атмосферу Земли органическими соединениями. Он допускал, что в противном случае жизнь на нашей планете возникла бы значительно позднее. Она еще не развилась бы при теперешнем возрасте Земли в 4,5 млрд. лет или же была бы представлена лишь самыми примитивными формами."Спустя несколько тысяч лет после своего образования Земля, вероятно, достаточно остыла, так что на ней образовалась довольно постоянная твердая кора. Однако в течение очень долгого времени температура этой коры была выше температуры кипения воды и конденсация воды происходила весьма медленно. Примитивная атмосфера содержала, возможно, очень мало кислорода или не содержала его вовсе... Почти весь углерод органических веществ и большая часть углерода, содержащегося сейчас в меле, доломитах и известняках, присутствовала в примитивной атмосфере в виде двуокиси углерода. Вероятно, значительная часть того азота, который сейчас находится в воздухе, была связана с металлами, образуя нитриды земной коры, так что под действием воды происходило непрерывное образование аммиака. Солнце светило, может быть, несколько ярче, чем сейчас, и поскольку атмосфера не содержала кислорода, ультрафиолетовые лучи не задерживались почти целиком слоем озона (аллотропная форма кислорода) в верхних слоях атмосферы и самим кислородом в более нижних ее слоях, как это происходит теперь. Они достигали поверхности суши и моря или, по крайней мере, облаков" **.Абиогенные синтезы. Холдейн указывал, что если пред-биотический мир был анаэробным, то любые появившиеся комплексные молекулы были более устойчивыми, чем в присутствии кислорода, и что тогда не мог еще существовать озон в атмосфере. Озон активно поглощает ультрафиолетовые лучи, и поэтому лишь небольшая их часть, входящая в состав солнечных лучей, проникает на Землю. До появления свободного кислорода в атмосфере ультрафиолетовые лучи могли проникать в больших количествах и быть мощным фактором для осуществления различных химических процессов.* J.B.S. Haldane. The origins of life. - "New biology", 1954 v 16, p. 12.** Дж. Б. с. Холдейн. Возникновение жизни, с. 300.
В работе "Возникновение жизни" (1929) изложен ход абиогенного синтеза биологически важных молекул в условиях первичной Земли. "Известно, что при действии ультрафиолетовых лучей на смесь воды, двуокиси углерода и аммиака возникает множество различных, органических соединений, в том числе сахара, а также, по-видимому, некоторых соединений, из которых образуются белки... Однако до того, как появилась жизнь, они, вероятно, накапливались, так что первичный океан достиг постепенно консистенции горячего жидкого бульона...".
Возникновение надмолекулярных систем из макромолекул. В работе "Происхождение жизни" (1957) Холдейн утверждает, что для жизни необходимы не только саморазмножающиеся молекулы, но и саморазмножающиеся системы таких молекул.
Мысль о постепенном усложнении организации носителей физической, химической и биологической форм движения материи Холдейн выразил в 1963 г. в своем выступлении на симпозиуме во Флориде:
"Физика в своих расчетах может предвидеть будущее. В физике практически установлено, что при объяснении свойств и поведения любой сложной физической системы, будь то атом, рассматриваемый как совокупность ядра и электронов, или молекула, рассматриваемая как совокупность атомов, на каждом этапе анализа мы сталкиваемся с новыми принципами организации. Теперь нам известны некоторые законы взаимодействия между атомами, между молекулами и т.д., изучаемые квантовой физикой, по для этого нам пришлось изучить сами молекулы. Я почти не сомневаюсь в наличии у живых систем значительно более сложных закономерностей того же типа, помогающих им сохранить свою структуру и функцию. Но я думаю, что мы еще недостаточно хорошо разбираемся в современной физике, более того я беру на себя смелость утверждать, что большинство биологов мыслят категориями физики 1920, а не 1963 г. ... При рассмотрении жизни в любой ее форме нужно всегда принимать во внимание, что в происходящих переходах участвуют квантовые закономерности" *.В работе "Возникновение жизни" (1929) это положение звучало несколько иначе:* Происхождение предбиологических систем, с. 70."Первые живые или полуживые объекты представляли собой, вероятно, большие молекулы, синтезировавшиеся под действием солнечных лучей и способные размножаться лишь в той исключительно благоприятной среде, в которой они возникли. Каждый из них, по-видимому, нуждался в целом ряде высоко-специализированных молекул для того, чтобы иметь возможность размножаться, а доступность этих молекул была делом случая".Для утверждения необходимости системной организации при возникновении жизни Холдейн приводит следующее сравнение:"Одноклеточные организмы, в том числе бактерии, которые прошлому поколению людей казались совсем простыми, устроены гораздо сложнее бактериофагов. Это организмы, иными словами, это уже целые системы, отдельные части которых действуют согласованно. Каждая часть специализирована для выполнения определенной химической функции и производит химические молекулы, необходимые для роста других частей. В результате клетка, как правило, может обходиться, потребляя молекулы нескольких типов, которые превращаются внутри нее в более сложные вещества, необходимые для роста ее частей".Там же Холдейн пишет:"Клетка состоит из многочисленных полуживых химических молекул, взвешенных в воде и заключенных в маслянистую пленку. В те времена, когда весь океан представлял собой огромную химическую лабораторию, условия для образования таких пленок были относительно благоприятными и тем не менее жизнь, возможно, находилась на «вирусной» стадии на протяжении многих лет, прежде чем подходящий набор элементарных единиц оказался собранным в первичной клетке. Этому событию, наверное, предшествовало множество неудач, однако первая удавшаяся клетка располагала обильной пищей и неизмеримыми преимуществами по сравнению со своими соперниками".В 1963 г. Холдейн отмечает, что у первых организмов весь обмен должен быть направлен в первую очередь на синтез нуклеиновой кислоты - матрицы для синтеза белка, катализирующего образование этой нуклеиновой кислоты.В работе 1957 г. Холдейн пришел к заключению, что при наличии синтезирующих ферментов и подходящих субстратов, по-видимому, может быть точно воспроизведена большая молекула ДНК. При наличии в растворе нуклеотидов, подходящего фермента и, по-видимому, источника свободной энергии вполне возможно самовоспроизведение. "Едва ли вызовет сомнение, - писал он, - что «лесами» для построения белков служат как ДНК, так и РНК" *.
* Цит. по кн.: "Происхождение жизни". Сб. "Планета Земля". М., ИЛ, 1961, с. 315.В лекции 1963 г. Холдейн вновь подчеркнул необходимость наличия источников свободной энергии. Самой важной является его мысль о необходимости макроэргических соединений (фосфатов) и одновременного присутствия необходимых для жизненных процессов низкомолекулярных органических соединений. В этой работе матрицей для синтеза белка он уже считал РНК, а не ДНК.В результате своих теоретических построений о первичности возникновения белков-ферментов или нуклеиновых кислот Холдейн оказался в замкнутом кругу. С одной стороны, для синтеза нуклеиновых кислот у первичных организмов необходимо наличие белков, катализирующих образование нуклеиновых кислот, а с другой - для синтеза белка нужна матрица - нуклеиновая кислота.
Чтобы выйти из этого заколдованного круга, Холдейн сделал два предположения. Первое: для образования нуклеотидов и их соединения между собой мог быть использован один неспецифический простейший фермент, являющийся, например, неспецифической фосфокиназой (переносчик фосфорной кислоты), который одновременно катализировал бы разные реакции (синтез нуклеотидов и их соединения в полимеры). Несмотря на такое предположение, Холдейн отлично понимал, что вероятность возникновения простейшего фермента типа рибонуклеазы (124 аминокислотных остатка) с необходимой специфической последовательностью аминокислот недопустимо ничтожна. Можно лишь думать о первичном возникновении активных полипептидов, значительно более мелких. Последнее подтверждается данными современной биохимии о том, что даже простые пептиды - участки белка, образующие активный центр, - могут обладать определенной ферментативной активностью.
Поэтому Холдейн выдвигает второе предположение. Не все комбинации, например в случае нуклеотидов РНК, являются одинаково вероятными по своим энергетическим возможностям. Некоторые конфигурации РНК могут быть значительно более вероятными, чем другие. Если бы среди них оказались биологически более целесообразные, то это резко повысило бы вероятность возникновения первичного жизненного субстрата (белка?). Холдейн предположил, что репликация РНК может быть вероятностным процессом, в результате которого могут образоваться различные варианты. Окончательная детерминация синтеза белков при усложнении первичной живой системы потребовала возникновения ДНК.
Первичные организмы. В работе "Возникновение жизни" (1929) Холдейн пишет, что
"первые предшественники жизни располагали пищей в больших количествах и у них не было соперников, с которыми им надо было вести борьбу за существование. Поскольку в примитивной атмосфере было очень мало кислорода, а может быть, и не было его вовсе, они добывали необходимую им для роста энергию не за счет окисления, а за счет брожения.Жизнь на других мирах. Вопроса о происхождении жизни Холдейн вскользь касается в лекциях, составивших позже книгу "Марксистская философия и науки" (1938). Он допускает, что первые организмы могли размножаться в инертной среде, но тут же указывает, что вирусы, нуждающиеся для размножения в организме-хозяине, могут потребовать от него размножения "подобно тому, как скороговорка требует непрестанного повторения". Холдейн замечает также, что при рассуждениях о химических процессах, связанных с происхождением жизни, можно впасть в ошибку, если ограничиваться понятиями в рамках современной химии (к этой мысли он возвращается еще раз в 1944-1945 гг.).Деятельность этих организмов должна была сильно зависеть, как это наблюдается у эмбрионов и бактерий, от ферментативных процессов, т.е. от добывания энергии с помощью каталитических реакций, протекающих на молекулярном уровне, когда эти процессы не связаны с окислительными процессами.
Возможно, что все ныне живущие организмы произошли от одного общего предка..., что жизнь возникла лишь однажды или, что более вероятно, потомки первого живого организма быстро продвинулись в своем развитии достаточно далеко, чтобы одолеть любых более поздних конкурентов, когда они появились на сцене.
Примитивные организмы использовали питательные вещества, содержащиеся в море, и некоторые из них начали осуществлять в своем собственном теле синтез, который прежде довольно нерегулярно осуществлялся под действием солнечного света; таким образом, эти организмы обеспечивали себе надежное снабжение пищей. Так возникли первые растения, обитавшие у поверхности океана и синтезировавшие пищу так же, как это делают их современные потомки.
По мнению многих биологов, мы, животные, происходим от этих растений. Среди молекул, составляющих тела, есть такие, которые по своей структуре сходны с хлорофиллом - зеленым пигментом, с помощью которого растения используют солнечный свет для своих нужд. Мы их используем для иных целей, нежели растения (например, для переноса кислорода) и мы, конечно, не знаем, являются ли они, так сказать, прямыми потомками хлорофилла или его родичами по боковой линии.
Все высказанные выше заключения носят спекулятивный характер. Они будут оставаться таковыми до тех пор, пока живые существа не будут синтезированы в биохимической лаборатории. Мы еще очень далеки от этой цели. Тем не менее я надеюсь дожить до той поры, когда будет синтезирован хотя бы один фермент. Вряд ли, впрочем, мне удастся дождаться того дня, когда будет синтезировано нечто живое, хотя бы в такой мере живое, как бактериофаг или вирус, и полагаю, что в течение ближайших столетий вряд ли удастся синтезировать какой-либо вполне самостоятельный организм. А до тех пор проблема происхождения жизни будет оставаться предметом спекуляций. Однако подобные спекуляции пе бесплодны, потому что они доступны экспериментальной проверке".
Холдейн начинает свое рассуждение с того, что в составе света, поступающего из Галактики, обнаружено "красное смещение". Если это - эффект Доплера и Вселенная непрерывно расширяется, тогда, считает Холдейн, можно воспользоваться выводом Милна * и допустить, что такое явление должно вызвать изменение во времени в процессе развития некоторых химических реакций, т.е. соотношения скоростей меж- и внутримолекулярных реакций должны подвергнуться изменениям. Холдейн высчитал, что в докембрийское время скорость, с которой энергия должна была высвобождаться из системы, напоминающей современную мышцу, была настолько мала, что наземное животное едва было в состоянии ползать.
* предельный случай космологической модели, ранее предложенной советским ученым А.А. Фридманом. Однако порочная методология привела Милна к религиозно-мистическим выводам о происхождении Вселенной. Холдейн-марксист принял активное участие в дискуссии по космологическим проблемам. В популярной статье в "Дейли Уоркер" он разъяснил суть дела с точки зрения методологии диалектического материализма, подчеркнув значение научных теорий и открытий для формирования материалистического мировоззрения в вопросах мироздания.В "Автонекрологе", вспоминая эпизод с гипотезой Милна, Холдейн со свойственным ему юмором заметил: "Я отважился сунуть нос даже в астрономию".Соотношение скоростей имеет непосредственное отношение и к диффузии, которая, по мнению Холдейна, является одним из главных факторов, влияющих на сохранение обмена веществ в клетках более простых организмов. Он допускает, что жизнь на клеточном уровне должна была выжидать наступления времени, когда изменившиеся химические процессы сделали бы возможным высвобождение соответствующего количества энергии, дабы сохранить требуемые условия внутренней среды.В работе "Происхождение жизни" (1957) Холдейн развивает мысль, что жизнь также могла возникнуть и на других мирах. Причем химические основы ее там могут быть совершенно отличными от наших. Например, он считал, что на внешних планетах Солнечной системы жизнь возможна при низких температурах. В основе ее могут лежать реакции, протекающие не в воде, а в жидком аммиаке.
Данные астрофизики о температуре на Венере и Марсе позволяют, по мнению Холдейна, предполагать наличие на них жизни типа земной. Однако отсутствие или незначительное содержание кислорода в атмосферах этих планет свидетельствует о том, что интенсивность фотосинтеза там не может быть сколько-нибудь значительной, чтобы способствовать возникновению животного мира.
"Если вообще возможно образование планет, - пишет Холдейн в этой работе, - сходных по составу с Землей, на определенном расстоянии от звезд, несколько мепыних по массе, чем Солнце, излучение которых придает им подходящую температуру, то на таких планетах жизнь могла бы поддерживаться в течение значительно более длительного времени. Возможно, что именно на таких звездах биохимическая и психическая эволюция могла бы достигнуть наибольшего расцвета... Астрономические исследования ближайших лет, вероятно, помогут дать ответ на все эти актуальные вопросы".Вклад Холдейна в разработку вопроса о происхождении жизни высоко оценил академик А.И. Опарин, посвятивший его памяти специальную статью, помещенную в мемориальном сборнике, вышедшем в Индии в 1965 г. Особенно он выделяет ту часть работы Холдейна 1929 г. "Возникновение жизни", где высказывается мысль о возможности существования "первичного бульона", подвергавшегося непрерывным изменениям.Со своей стороны Холдейн рассматривал работы А.И. Опарина как весьма важные для развития теории происхождения жизни на Земле. Он писал:
"А. Опарина и меня можно рассматривать как древнейших представителей той науки, которая занимается вопросом происхождения жизни на Земле. Однако между нами существенная разница. В то время как я не очень глубоко разбираюсь в этом вопросе, Опарин посвятил ему всю свою жизнь.По мнению Дж. Бернала, гипотеза Опарина-Холдейна вывела проблему происхождения жизни из сферы чистых спекуляций, началась эра поисковых экспериментов в области синтеза **.Нет никакого сомнения в том, что приоритет принадлежит не мне, а профессору Опарину. К своему стыду, я не читал его ранней работы и поэтому не могу ничего сказать. Однако, насколько я помню, мои первые печатные работы появились в 1927 г., а его - в 1924 г. В моей маленькой статье вряд ли можно найти что-либо новое, чего не было бы в его книге. Речь может быть только о плагиате" *.
* J.В.S. Haldane. Data needed for a blueprint of the first organism. - In: "The origins of prebiological systems". New York, Academic Press, 1965, p. 11.** Происхождение предбиологических систем, с. 77.
Биохимия наследственности и биохимическая теория эволюции
Зарождение биохимической генетики следует датировать началом 30-х годов, когда она еще называлась "физиологической генетикой". В ее основу было положено несколько направлений исследований, прежде всего опыты по анализу развития пигментации путем трансплантации имагинальных дисков глаза у насекомых (Бидл и Эфрусси, 1936) и работы по изучению биохимии наследования окраски цветков у примулы, выполненные Дж. Холдейном и Д. Де Уинтоном (1930-1935).
Исследования Бидла и Эфрусси по своему идейному замыслу и технике выполнения явились прямым продолжением методик, заимствованных из экспериментальной эмбриологии, и состояли в основном в трапсплаптации зачатков органов. Полученные результаты убедительно говорили в пользу биохимической природы генной ре гуляции и подсказали пути поисков соединений, участвующих в синтезе пигментов глаза.
В скором времени были уже сформулированы основные положения биохимической генетики на основе достижений химии ферментов, в частности опытов с классическим объектом - нейроспорой, выполненных Биллом, Татумом и их сотрудниками. Последним удалось получить много мутаций, нарушающих пути синтеза аминокислот, витаминов и. других соединений, и установить основные взаимосвязи этих процессов у нейроспоры. Они осуществили анализ ферментативных систем, участвующих в этих реакциях. Хотя окончательные формулировки положений биохимической генетики были даны Бидлом и Татумом на основе работ по дрозофиле и нейроспоре, первые этапы развития пауки связаны с исследованиями заболеваний человека (А.Е. Гаррод, 1902; Л. Кэно, 1903) и пигментов цветков (Дж. Холдейн).
Современные историки генетики Е.А. Карлсон, Л.С. Данн, Дж. Бидл и другие единодушно отмечают, что еще в период работы в лаборатории Гопкинса при изучении врожденных парушений обмепа веществ у человека Холдейн был одним из первых, кто обратил внимание на забытое исследование английского ученого А.Е. Гаррода, установившего в 1902 г. связь между действием гена и фермента *.
* А.Е. Garrod. Inborn errors of metabolism. Oxford, 1909.Е.А. Карлсон пишет: "Следует особо отметить заслуги перед биохимической генетикой Джона Б.С. Холдейна, как выдающегося теоретика. Он популяризировал работу Гаррода «О врожденных нарушениях» еще до ссылок на пее в исследованиях Дж. Бидла и Э. Татума. Он попытался связать генетику с биохимией в своей монографии «Биохимия генетики». К сожалению, Холдейн редко доносил свои теоретические построения до лабораторного воплощения, и разработку этого вопроса биохимической генетики в 1940 г. со всей справедливостью следует приписать Бидлу и Татуму" *.* E.A. Carlson. The gene: A critical history. Philadelphia, 1966.Сам же Бидл считает, что"книга Гаррода знаменует начало нового этапа - создания современной биохимической и медицинской генетики. Хотя работа Гаррода упомянута в работе Бэтсона (1913) и в его собственной книге, переизданной в 1923 г., она как-то затерялась на полках научных библиотек. Биохимики не интересовались генетикой, а генетики - биохимией. Лишь в середине 30-х годов в связи с изучением химии растительных пигментов были разработаны теории, весьма близкие к построениям Гаррода. Его спас из забвения английский генетик Джон Б.С. Холдейн, который в 1942 г. отметил, что открытие связи гена с ферментом, которое выдается за новинку, было предвосхищено еще тридцать лет назад Гарродом" *.Холдейн был скорее теоретиком, чем экспериментатором в различных областях генетики (как экспериментатор он известен в основном в качестве физиолога, ставившего опыты на самом себе), его вклад в биохимическую генетику следует рассматривать в плане теории, главным образом в истолковании биохимической природы действия генов. Этому вопросу в основном посвящены его труды: "Новые направления в генетике" (1941), "Биохимия генетики" (1954) * и некоторые разделы "Энзимов" (1930).* G. and M. Beadle. The language of life. New York, 1966, p. 130.* По традиции, установившейся в пашей научной литературе, работу Холдейпа "The biochemistry of genetics" ошибочно переводят "Биохимическая генетика", что несколько обедняет смысл и сужает круг проблем, стоящих перед этим ведущим разделом современной генетики, ориентируя скорее на изучение генетики биохимических признаков. Монографию Холдейна следует именовать "Биохимия генетики" или "Биохимия наследственности", поскольку она посвящена анализу природы биохимического механизма действия генов.Когда в 1928 г., после кончины Бэтсона, Холдейп возглавил Институт садовых культур, по его инициативе были начаты исследования пигментов, направленные на систематическое изучение биохимической основы действия генов. В качестве объекта исследования избрали лепестки цветков, имея в виду бесконечные вариации их окраски, а также потому, что в отличие от большинства пигментов животных пигменты лепестков можно было получить в относительно больших количествах и они отличались высокой растворимостью. Это обстоятельство облегчало работу с пигментами в лабораторных условиях.Из-за отсутствия средств программа работ оказалась неосуществленной; однако в 30-х годах интерес к проблеме не угас, и вскоре группа молодых ученых опубликовала несколько сообщений по этому вопросу *. Холдейн тоже участвовал в работах подобного рода, ему, например, удалось показать, что рН клеточного сока у растений контролируется генетическими механизмами.
* См., например: R. Scott-Moncrieff. - "J. Genetics", 1936, v. 32, p. 117 и др.К концу 30-х годов химия пигментов явилась одним из наиболее разработанных разделов биохимии растений. К этому времени уже была установлена структура, изменчивость и основные типы пигментов, а также гены, контролирующие состав различных пигментов, процесс образования и торможения синтеза коферментов, условия окисления и метилирования пигментов, вариации рН клеточного сока у ряда видов растений. Были поставлены опыты по изучению роли различных углеводов в формировании пигментов.Итоги исследований Холдейна и его сотрудников в области генетических механизмов, контролирующих пигментацию цветков, были изложены в цикле лекций, объединенных в монографию "Новые направления в генетике" (1941). В книге много внимания было уделено проблеме гомологии генов у разных организмов, проблеме, которую так успешно (правда, в другом аспекте) разрабатывал Н.И. Вавилов, в частности в своей классической работе "Закон гомологических рядов в наследственной изменчивости" (1920). Если Н.И. Вавилов изучал главным образом направления изменчивости признаков, то Холдейн сосредоточился на исследовании отдельных генов в аспекте их влияния на элементарные биохимические особенности организма. Ему удалось выявить сходство в действии аналогичных генов у разных видов растений и даже подобие в локализации их в хромосомах, что он исследовал еще в 1930-1935 гг. совместно с Де Уинтом.
Проблема гомологии генов имеет общебиологическое значение, так как решает вопрос, происходят ли сходные гены разных видов от одного и того же "гена-предшественника", или они возникли независимо, в ответ на одинаковые требования среды (подобно конвергенции признаков). При анализе закономерностей действия генов Холдейн придавал большое значение времени проявления генов в развитии организма. В работе "Время действия генов и их связь с некоторыми вопросами эволюции" (1932) на основании изучения генетического материала, полученного на растительных и животных объектах, Холдейн приходит к трем важным выводам:
1. Гены можно классифицировать в соответствии с временем их действия. Они могут действовать не только в диплоидных зиготах, эндоспермах и гаметах, несущих гены, но и в течение периода, превышающего жизненный цикл организма *.Сопоставляя разные направления исследований Холдейна - генный контроль пигментации, проблему доминирования, гомологию генов - необходимо подчеркнуть взаимосвязь, казалось бы, совершенно разных проблем: биохимической природы и механизма действия генов, роли и принципов изменчивости генов в эволюции и, наконец, их участия в индивидуальном развитии организма. Все эти проблемы были в центре интересов Холдейна и связаны с основным вопросом о химической природе и механизме действия гена.* В частности, генотип матери может отразиться на фенотипе потомства.2. Предполагается, что изменение времени действия генов является важным фактором в эволюции и что некоторые случаи ортогенеза, включая дегенерацию, можно объяснить временем действия гена.3. В организме, подверженном метаморфозам, действие гена, определяющего успех адаптации, зависит от ограниченного времени его действия.
В монографии 1941 г. "Новые направления в генетике" Холдейн проводит аналогию между развитием ферментативных реакций и событиями на клеточном и субклеточном уровне и замечает:
"Если за десять лет работы под руководством такого выдающегося биохимика, как Гопкинс, я убедился лишь в том, что биохимическое истолкование биологических явлений является более основательным, нежели морфологический подход, то я все же что-то усвоил и в чем-то преуспел".Далее он пишет:"Так же как в случае с превращениями ферментов, геп превращается в одну из своих форм - рецессивный аллеломорф и более не выполняет ¦свою первоначальную функцию или же выполняет ее значительно медленнее. При биохимическом подходе к изучению природы гена можно получить всестороннюю информацию о функции каждого гена. Но поскольку каждая клетка, по-видимому, содержит несколько тысяч генов, то потребуется самое тщательное и всестороннее изучение биохимии клетки, и это дело будущих поколений биохимиков и генетиков. На этом пути наука обогатится сведениями и в области физиологии развития, что будет бесценным приобретением" *.Основную гипотезу, известную как закон "один ген - один фермент", Холдейн сформулировал еще в 1920 г. Он был уверен и часто подчеркивал, что она справедлива в отношении всех организмов, в том числе и человека.* J.В.S. Haldane. New paths in genetics. London, 1941, p. 24.Однако идея эта была высказана не только Холдейном, Независимо от него и почти одновременно с ним С. Райт и Р. Гольдшмидт * пришли к аналогичному выводу.
Холдейн писал, что эта мысль возникла у него под. влиянием работы Л. Кэно, который впервые исследовал генетические особенности пигментации у мышей и пришел к выводу о взаимодействии и контролирующем влиянии генов, участвующих в формировании окраски покровов **.* См. Haldane and modern biology. Baltimore, 1968, p. 45.Холдейн рассматривал гены как органы клетки и считал, что они выполняют определенную функцию. Он допускал, что гены есть не что иное, как клеточные катализаторы, весьма напоминающие по своим свойствам ферменты, и что они продуцируют определенное количество специфических ферментов; аллели же образуют ферменты, лишь количественно отличающиеся от специфических ферментов, которые продуцируют нормальные гены. Эта теория весьма близка к теории действия гена, предложенной в 1927 г. Р. Гольдшмидтом в том отношении, что в обоих случаях авторы допускают, что ген и его продукт идентичны и что мутации - это всего лишь изменения в количестве продуцируемого фермента. Точно так же идентифицировал гены и их продукты советский генетик Н. К. Кольцов *.* J.B.S. Haldane. The biochemistry of genetics. London, 1960.
* См. Н.К. Кольцов. Организация клетки. М., 1936, с. 585, 623.Пожалуй, наиболее богатый материал в пользу такого представления давали работы по иммуногенетике, по изучению влияния генов на синтез антигенов, прежде всего в крови животных и особенно человека. К вопросу о наследовании антигенов Холдейн возвращался неоднократно,, например в 1935 г., а потом снова в 1953-1954 гг. *. Но в то же время неясность проблемы, неочевидность идентичности гена и его продукта заставляли Холдейна рассматривать и другие возможности, модифицировать гипотезу. Так, в 1935 г. он развил теорию "один ген - один фермент", показав регулирование деятельности специфических ферментов генами.* J.B.S. Haldane. Contribution de la genetique a la solution de quelques problemes physiologiques. Reunion Pleniere de la Soc. de Biologie. Paris, 1935; J.B.S. Haldane. The genetics of somу biochemical abnormalities. Lecture on the scientific basis of medicine, 1953-1954, v. Ill, p. 41.Поскольку Холдейн по складу своего мышления был абсолютно свободен от догматизма, он неоднократно пересматривал эту проблему (которая была предметом его постоянного интереса) и вносил в нее поправки. В более поздние годы он допускал, что первичный генный продукт отличается от самого гена, и затем отказался от мнения, что аллели одного и того же гена отличны друг от друга лишь количественно. В конце 30-х годов Холдейн утверждал, что мутантные гены отличаются от исходных генов (в пределах одного локуса) не только количественно, но и качественно. Он считал, что отдельный "наследственный признак" может развиваться лишь при паличии множества генов (следовательно, должно быть, по крайней мере, два доминантных гена или более) и был убежден в том, что действие заданного набора генов зависит от внешней среды.Холдейн отмечает, что уже во втором десятилетии XX в. Хагедурн (Hagedoorn) рассматривал гены как субстанцию, определяющую ход внутриклеточных процессов. "В настоящее время, - пишет он, - мы все больше осознаем, что намного приблизимся к попиманию истинного развития событий, если будем рассматривать ген как субстанцию, ведающую координацией процессов, а не как «наследственную единицу»", и далее подчеркивает, что его взгляд является "лишь одной из ступенек на бесконечном пути к познанию истины" *.
* J.B.S. Haldane. New paths in genetics. London, 1941, p. 21.Особое внимание Холдейн уделял вопросу пространственного расположения генов в клетке и связям его с другими генами. Он допускал, что среди генов существует функциональная специализация - от одних зависит выработка группы гормонов (или гормона), от других - окислительно-восстановительные процессы и т.д. Холдейн рассматривал ген как субстанцию белковой природы, которая по своему размеру скорее всего приближается к величине белковой молекулы, но не исключал, что ген, подобно вирусу, является нуклеопротеидом.По мнению Холдейна, функция гена, его координирующая деятельность должна быть охарактеризована в химических понятиях. Весьма трудно допустить, что ген непосредственно управляет окислительными процессами, т.е. участвует в них как каталитический агент, скоpee следует предположить, что он играет в координации опосредствующую роль. Холдейн даже высказывал мысль, что регуляторная функция гена (ген - окислительный процесс) может быть и многоступенчатой природы ("многозвеньевой"), напоминая систему "причинных ступенек", наблюдаемую между событиями, разыгрывающимися между моментом активности нервного центра и сокращением группы мышц.
Когда в 1940-1950-х годах распространилось мнение о ведущей роли нуклеиновых кислот в наследственности, Холдейн выступил с гипотезой, которая, как мы теперь знаем, оказалась в принципе верной. В докладе на VI коллоквиуме Общества физиологической химии "Значение макромолекул для эволюции и дифференцировки" (1956) он говорил, что имеется два типа макромолекул. Первый - линейные (неветвистые) макромолекулы, как ДНК или протамины. Только такие молекулы способны копироваться, сохраняя мутационные изменения. Второй тип макромолекул - нелинейные, как, например, инсулин, вазопрессин пли окситоцин. Они не копируются.
Правильность этой гипотезы заключается в том, что, действительно, копироваться при редупликации могут только линейные макромолекулы ДНК или РНК (у вирусов). Ошибка Холдейна заключалась в том, что в группу способных к копированию макромолекул он зачислил и некоторые белки. Между тем теперь известно, что перекодирование белок-белок или белок-нуклеиновые кислоты невозможно или, во всяком случае, в живой природе этого не происходит *. Теперь это принимается как "основная догма молекулярной биологии".
* См. F. Crick. The central dogma of molecular biology. - "Nature", 1970, v. 227, p. 561-563.Ошибка Холдейна была связана с тем, что он не сразу отказался от признания непосредственной роли белков в пострении гена, не сразу принял положение о том, что основой гена, носителем содержащейся в нем наследственной информации является исключительно ДНК (РНК у некоторых вирусов). Вероятно, такое неприятие исключительной роли ДНК объясняется тем, что, как уже говорилось, первоначально Холдейн идентифицировал ген и его продукт - белок (фермент). Однако основная идея Холдейна "один ген - один фермент" оказалась правильной и в высшей степени плодотворной.Холдейн не сделал в биохимической генетике принципиально важных фактических открытий. И все же он активно участвовал в становлении нового биохимического направления, ввел в генетику новую методологию. В 1941 г. он писал:
"У меня нет никакого намерения умалять выдающиеся достижения формальной генетики в прошлом. Такие деятели науки, как Мендель, Бэтсон, де Фриз и Морган внесли существенный вклад в развитие генетики, подобно Везалию или Фабрицию. И тем не менее в настоящее время исследования в области чисто формальной генетики уже не имеют первостепенного значения *. Современный генетик находится в несколько затруднительном положении. Он должен быть хотя бы поверхностно знаком с анатомией, цитологией и матема тикой. Он должен что-то смыслить в таксономии, физике и даже психологии. И если он не ученик Гопкинса, он порой может забыть, что биохимия - это ключ к решению любых вопросов фундаментального значения для органического мира" **.Самым существенным вкладом Холдейна в биохимическую генетику следует считать его методологический подход к изучению природы гена и его действия. Он стоял на правильном пути, когда неоднократно подчеркивал, что этот вопрос может быть решен только в понятиях биохимии. В последние годы наука значительно приблизилась к углубленному познанию сущности гена и его действия, уже получены гены в изолированном виде. В 1920-1930 гг. Холдейн был, пожалуй, одним из очень немногих, ставивших со всей остротой вопрос о необходимости вести интенсивный научный поиск сопряжепных связей между геном и ферментом. Он явился, как отмечают многие его современники, одним из пионеров, объединивших биохимию с генетикой. Именно в ту пору, когда он работал над своим большим трудом "Биохимия генетики" - кстати сказать, одной из первых книг в этой области в мировой литературе - были сделаны важнейшие открытия в молекулярной генетике, заключавшие в себе ответ на многие вопросы, поставленные в этой монографии. В 1954 г. Холдейн одобрительно отозвался о грустном "факторе старения" сказав: "Так уж получается, как часто в науке бывает, что от момента написания книги до публикации материал и мысли устаревают".* Дальнейшее развитие науки о наследственности по пути молекулярной генетики показало принципиальную правильность этого мнения Холдейна.** J.B.S. Haldane. New paths in genetics. London, 1941, p. 46.
С середины 50-х годов из-за отсутствия необходимых условий для лабораторных исследований Холдейн уже не участвовал в постановке экспериментов. Однако он всегда был в курсе новейших направлений и открытий, откликаясь на них, выступая в качестве компетентного и авторитетного ученого.
Его гипотезы имели большое значение для разных областей биохимической генетики, а не только для проблемы природы и механизма действия генов, о чем говорилось выше. Эрнст Каспари, например, отмечает *, что, быть может, самым значительным вкладом Холдейна в биохимическую генетику в последний его период была высказанная им гипотеза о том, что наиболее примитивной формой жизни должны были быть ауксотрофы (организмы, не способные расти на минимальной питательной среде, так как они не могут синтезировать какие-либо необходимые органические вещества; для их роста требуется поступление этих веществ извне). По мнению Холдейна, первичные ауксотрофные организмы могли добывать энергию из органических соединений. Последние в свою очередь могли образовываться под действием солнечных лучей в атмосфере, лишенной молекулярного кислорода. Эти теоретические построения являются основой современной биохимической теории эволюции. Они получили дальнейшее развитие в работах А.И. Опарина и могли быть подвергнуты экспериментальной проверке.
* Haldane and modern biology. Baltimore, 1968, p. 49.Очень точные слова для характеристики могучего таланта Холдейна нашел польский ученый Гаевский:"Холдейн, без сомнения, один из любопытпейших умов среди современных биологов... Из огромного количества экспериментального материала, поставляемого ежегодно тысячами биологов, он сумел отобрать самое существенное и построить общие теории и гипотезы. Это связано с его удивительным талантом постановки научных проблем под новым углом зрения. Причем изложепие проблем всегда отличается предельной четкостью и точностью. Это достоинство объясняется также его склонностью к широким математическим обобщениям процессов жизнедеятельности в их теснейшей взаимосвязи и взаимоопределяемости" *.* См. W. Gajewski. Предисловие к польскому изданию книги Дж. Холдейна "Биохимия генетики". Варшава, 1960.Математика
"Математика - муза моя", - говорил Холдейн. Для математического склада его мышления характерен, например, такой случай. "Однажды, - рассказывает профессор Д. Льюис, - я застал его одного в кабинете в каком-то необыкновенно благодушном настроении. Вдруг Холдейн сказал: «Сегодпя у меня день рождения, но самое интересное, что мой возраст равен квадратному корню из текущего года»" . Это было 5 ноября 1936 г., когда ему исполнилось 44 года.
Математика - язык науки, и, так же как нельзя представить себе, скажем, поэта, не владеющего литературным языком, в наше время не может быть ученого, не знакомого с языком математики. Холдейн не только блестяще владел "математическим языком", но и успешно совершенствовал его. Он в каком-то отношении олицетворял собой лучшие традиции ученых прошлого, сочетая в себе необычайно острое абстрактное мышление с поразительным умением не отрываться от конкретного смысла исследуемой проблемы.
Холдейн часто с благодарностью вспоминал о своих учителях в области математики и биометрии: Френсисе Гальтоне, Карле Пирсоне и Г.X. Харди, лекции которого по высшей математике в Оксфорде и Кембридже были значительным явлением для того времени.

Г.Х. Харди
Математические идеи, перенятые Холдейном у Харди, получили свое развитие и применение в области биологии и многих как смежных, так и далеко от нее отстоящих областей знания. После смерти своего учителя он писал: "Я скорблю об ученом, дорогом мне человеке, математические работы которого вызывают у меня переживания, подобные тем, что испытывают иные, слушая произведения музыкальной классики" *.
* J.B.S. Haldane. Everything has a history. London, 1951, p. 242.С такой же теплотой Холдейп отзывался о Пирсоне, несмотря на критическое отношение к его философским взглядам. В лекции, посвященной столетию со дня рождения К. Пирсона, Холдейн отмечал большие заслуги этого ученого, который разработал статистические таблицы и изучил вопросы, связанные с прикладным использованием теории вероятностей, причем исследовал также и область применения графических методов для оценки функций распределения. Эти новые для того времени методы были посвящены обоснованию математической теории эволюции. Холдейн подчеркивает, что не будет преувеличением сказать, что последующее успешное развитие теоретических основ математической статистики в значительной степени обязано усилиям Пирсона в этой области в десятилетие 1893-1903 гг. В эти же годы, по инициативе Пирсона, стал издаваться журнал "Биометрия". Холдейн считает, что зачатки многих дальнейших разработок в области статистики можно обнаружить в работах Пирсона: "Не следует забывать, что если нам, последователям Пирсона, и удалось придать теории математической статистики относительную доступность, даже принимая во внимание тот факт, что значительная часть математических разработок, предложенных им, уже утратила свою актуальность, то всеми этими успехами мы все же обязаны тому, что продвигались по пути, проложенному титаном мысли" *.* J.B.S. Haldane. Karl Pearson. - "Biometrika", 1957, v. 44, p. 303.
Карл Пирсон
Изучение крайне многообразного научного наследия Холдейна убедительно говорит о том, что вне математики и без математики оно немыслимо. Ученый сам весьма ярко и эмоционально говорит об этой склонности своей натуры: "Я принадлежу к числу тех, кто находит истинное эстетическое наслаждение в занятиях математикой, независимо от ее прикладного значения. Я сознаю, что этого недостаточно, но отдаю себе отчет в том, что люди, извлекающие наибольшее наслаждение из любимого предмета, могут, по всей вероятности, максимально обогатить и развит.ь этот предмет" *.
* J.B.S. Haldane. Everything has a history. London, 1951, p. 242.Высшее назначение математики Холдейн видит именно в том, чтобы с ее помощью находить скрытый порядок в хаосе, окружающем человека. Для пего абстрактные математические теории всегда имеют непосредственное отношение к описанию явлений природы."Занятие дифференциальным исчислением является само по себе истинно захватывающим делом. Мне вспоминается, как в феврале 1917 г., после ранения на европейском театре военных действий, я среди прочих офицеров был переправлен на пароходе вниз по реке Тигр, в Месопотамии. Лежа на носилках, я читал математическую книгу о векторах, а рядом лежащий офицер углубился в книгу по дифференциальному исчислению. Мы оба предпочитали такую литературу художественной, и опа служила нам наилучшим болеутоляющим средством. Некоторые разделы математики так изящны и прекрасны, что могут быть поставлены в один ряд с величественными образцами поэзии или живописи. Дифференциальное исчисление прекрасно, но совсем не потому, что является продуктом «чистой мысли», скорее наоборот, оно возникло, как вспомогательное средство для расчета движений звезд и полета пушечных ядер. По своей красоте опо вполне сопоставимо с изящно действующим машинным устройством".И далее: "Наука почти всегда имеет количественное выражение. Она имеет дело с измерениями, весовыми показателями и счетом" *.* J.B.S. Haldane. A banned broadcast and other essays. London, 1946, p. 116.Занятия Холдейна прикладной математикой сыграли поистине неоценимую роль в деле обороноспособности Англии. Как уже говорилось, 29 октября 1938 г. в журнале "Nature" появилась статья Холдейна "Математика ПВО" с расчетами вероятности и степени защищенности населения больших городов от возможности налета гитлеровской авиации.В период 1941-1945 гг. Холдейн по заданию командования военно-воздушных и сухопутных сил и министерства самолетостроения Англии решил ряд важных проблем оборонного значения на основе методов статистического анализа. Им, в частности, была разработана система, благодаря которой служба береговой обороны могла вести поиск и устанавливать местонахождение вражеских подлодок, действующих на ближних подступах к Британским островам. Он участвовал в разработке математических основ для создания специальной авиационной техники, с помощью которой разведывательная авиация сбрасывала на намеченные для бомбометания объекты специальную аппаратуру, обеспечивающую наведение на цель самолетов бомбардировочной авиации. Позднее Холдейн разработал математическую основу методов действия ПВО Англии по уничтожению в воздухе беспилотных аппаратов противника.
Эти математические работы, поставленные на службу обороны, в частности "Статистическое исследование потерь вооруженных сил", подготовленное по заданию армейского командования Великобритании были не чем иным, как продолжением, а порой дальнейшим развитием исследований в области генетики человека. Это, в частности, четко прослеживается в той части работы Холдейна, где подчеркивается связь между статистической вероятностью поражения целей в некоторых районах и статистической вероятностью наследственности.
В ряде работ Холдейн рассматривал случаи возможного применения математики к генетическим исследованиям и к изучению эволюционного процесса. В статье 1949 г. "К вопросу о количественных измерениях темпов эволюции" он дает высокую оценку основным положениям книги Симпсона *, посвященным геологическому прошлому состава диатомовых.
* G.G. Simpson. Rates of evolution in animals. - In: "Genetics paleontology and Evolution". Princeton, 1949.В последний приезд из Индии в Лондон Холдейн в письме к Н.У. Пири от 18 февраля 1964 г. писал по этому поводу:"Дорогой Билл,Зорко следя за успехами наук по самому широкому фронту, Холдейн, по мнению его современников, был одним из самых осведомленных ученых своего времени. Его дар к экстраполяциям в области математики был в значительной степепи продиктован глубоким убеждением в том, что на нынешнем этапе развития естествознания необходимо ломать рамки узкой специализации, наносящей вред объединению наук о природе. Он полагал, что наиболее надежным средством для этой цели должен быть набор математических инструментов, посредством которых ученому предстоит испытать их действенность на цифровом материале.Мной создано два новых понятия:
(1) Морганида. Единица измерения длины хромосомы по карте. Наиболее эффективной единицей следует считать «сантиморганида» (сокращенно «сМ»). См.: «Journal of Genetics», 1921, №1. Единицей «сМ» широко пользуется Г.С. Картер в ряде работ, посвященных сцеплению у мышей, относящихся к 1955 г.
(2) Дарвин. Признак, изменяющий свою среднюю величину в е раз за период длиной в миллион лет или же в 1,001 раза за тысячу лет. При этом допускается, что признак изменяется со скоростью в 1 дарвин. Средняя скорость (темп) эволюции линейных размеров зубов лошади, начиная с эоцена, примерно равна 40 миллидарвинов. Этот термин введен мной. Высота черепа у человека, видимо, увеличивалась со скоростью около 1 дарвина в период плейстоцена. Но длина тела динозавра изменялась со скоростью около 40 миллидарвинов (несомненно, скачкообразно). Соотношение длины (фактор формы), наблюдаемое в размерах зубов лошади, изменялось при величине около 20 миллидарвинов. Симпсон в I гл. «Основные черты эволюции» пересмотрел предложенные мною величины, пользуясь более совершенной шкалой времени *. Один бельгийский палеонтолог, не могу вспомнить его имени, пользовался миллидарвинами".
* "Он ссылается на мою статью. Мне помнится, что он пользуется терминами «килодарвин» и «мегадарвин». Последнее вполне применимо к некоторым домашним животным, в отличие от скачков в менделевском понимании. Что касается темпа эволюции нуммулитов [ископаемый вид фораминиферы] то их, по-видимому, можно измерять в килодарвинах".Руководствуясь этими методологическими соображениями, Холдейн предложил позаимствовать из астрофизики единицу "декс", введенную Ч.У. Алланом. "Декс" (от английского "decimal exponent") есть показатель степени при 10 и преобразует впереди стоящее число в десятичный антилогарифм. Он способствует более лаконичному написанию больших чисел, например, вместо 1039 - 39 дексов. Этой единицей удобно пользоваться для количественных характеристик в различных областях биологии. К примеру: организм человека состоит из 14 дексов клеток, или: соотношение длины и ширины нервного волокна может превышать 6 дексов. Холдейн убедительно показывает, что "дексы" могут найти себе широкое применение и в социологических исследованиях. При этом он отмечает, что "биологам нужно приучать себя к тому, чтобы быстро и оперативно мыслить категориями больших чисел, как это давно имеет место среди физиков, химиков, астрономов и геологов. При работе с малыми числами также весьма целесообразно пользоваться этой единицей, поскольку это способствует выработке точности мышления у исследователя" *.
* J.B.S. Haldane. "Dex" or "order of magnitude". - "Nature", 1960, v. 187, p. 879.Несмотря на все сказанное, чисто математические работы Холдейна (в общей сложности их было 30) также представляют собой серьезный и вполне самостоятельный интерес. В ходе изучения генетики популяций Холдейну пришлось не только столкнуться, но и серьезно заняться теорией уравнений в конечных разностях. Именно этому разделу математики суждено было стать основой современных методов вычислений на ЭВМ. Одним из блестящих математических достижений Холдейна в этой области был найденный им метод решения нелинейного разностного уравнения вида *y(u, Du) = 0, где Du = u(x+1) - u(x).
Этот метод был впоследствии включен в классическую монографию "Исчисление конечных разностей" английского математика Милн-Томсона и был им охарактеризован как "элегантный".
* "Proceedings of Cambridge Philosophical Society", 1932, v. 28, p. 234.Профессор М.С. Бартлетт, близко знавший Холдейна, вспоминает:"Виртуозно владея математической техникой в области классической статистики, Холдейн очень часто на досуге любил заниматься (и не всегда безошибочно!) разработкой некоторых алгебраических деталей, в частности расчетом нечетных моментов статистических распределений. Эти занятия привели его к разработке ряда интересных методов в математической статистике, связанных с вопросами применимости теории крупномасштабной выборки к встречающимся на практике реальным условиям ограниченного числа опытов.Эта серия исследований включала: работы, начатые еще в 1937 г., в которых Холдейн провел вычисления точных значений моментов так называемого c-распределе-ния, широко используемого при статистических оценках; работы, начатые Холдейном в 1951 г., в которых даны оценки среднеквадратичных отклонений и других моментов при определении эффективных значений параметров по методу наибольшего правдоподобия; и наконец, цикл исследований, предпринятых в 1938 г. с целью дать приближенное аналитическое описание целого ряда встречающихся на практике законов распределения случайных величии" *.
* Цит. по кн.: N. W. Pirie. J.B.S. Haldane. - Biographical memories of fellows of the Royal Society, 1966, v. 12, p. 233.
На лекции по биометрии в Лондонском университете (1953)
![]()