том 73, № 7, с. 614-621 (2003)
|
|
ВЕСТНИК РОССИЙСКОЙ АКАДЕМИИ
НАУК
том 73, № 7, с. 614-621 (2003) |
© Э.К. Хуснутдинова
ЭТНОГЕНОМИКА И ГЕНЕТИЧЕСКАЯ
ИСТОРИЯ НАРОДОВ ВОСТОЧНОЙ ЕВРОПЫЭ. К. Хуснутдинова
Хуснутдинова Эльза Камилевна - член-корреспондент Академии наук Республики Башкортостан,
заведующая отделом Института биохимии и генетики Уфимского научного центра РАН.Эволюция человеческих сообществ (популяций), их происхождение, родство, историческое развитие всегда были в центре внимания многих наук. Для решения этих проблем необходимо исследовать множество признаков в большом числе популяций и этнотерриториальных групп. В качестве таких признаков можно использовать вариабельность структуры биополимеров (белки. нуклеиновые кислоты). Раньше основное внимание уделяли полиморфным белкам. Подлинный переворот в исследованиях произошел при появлении нового типа маркеров, основанных на геномных ДНК.
Геном человека, состоящий примерно из 3 млрд. нуклеотидных пар, расшифрован почти полностью. Однако само по себе близкое завершение этой гигантского по замыслу и грандиозного по реализации международного научного проекта отнюдь не означает, что процесс познания генома завершен. Уже сейчас очевидно, что не существует какого-то "усредненного" генома человека: каждый геном, как и каждый человек, сугубо индивидуален. Эта индивидуальность проявляется на уровне не только отдельной личности, но и этнических групп, отдельных сообществ и рас.
Различия между двумя людьми на уровне ДНК составляют в среднем один нуклеотид на тысячу. Именно этим обусловливаются наследственные индивидуальные особенности каждого человека. Заметим, что различия между ДНК человека и шимпанзе - его ближайшего сородича в животном мире - на порядок больше.
ОСНОВНОЙ ИНСТРУМЕНТ ЭТНОГЕНОМИКИ
Для исследований геномов людей используют разные типы ДНК-маркеров: расположенные на парных хромосомах (аутосомные), на митохондриальной ДНК и на непарной Y-хромосоме. Маркеры на парных хромосомах наследуются по обеим - женской и мужской - линиям, в них представлена подавляющая часть генома человека. Таким образом, у нас в руках оказались маркеры, позволяющие изучать сочетанную изменчивость, одновременно привносимую и с отцовской, и с материнской стороны. Все маркеры ДНК характеризуют сообщества в целом, не выделяя генетического вклада каждого из полов. Используя определенные типы полиморфизма ДНК, можно оценить те или иные временные события, происходившие в истории данной популяции.
Особую роль играют маркеры митохондриальной ДНК (мтДНК) и ДНК Y-хромосомы, поскольку они помогают проследить генетическую историю человечества отдельно по женской и мужской линиям. Митохондриальная ДНК передается потомкам только от матери, так как митохондрии находятся в цитоплазме клетки, а цитоплазма потомка (зиготы) образуется за счет цитоплазмы материнской яйцеклетки. Если два человека имеют общего предка женского пола, то по различиям их мтДНК можно судить о том, сколько поколений отделяет их от жившей столетия или тысячелетия назад общей пра-... прабабушки. Аналогично изучение ДНК Y-хромосомы позволяет проследить эволюционные траектории по отцовской линии, поскольку Y-хромосома передается только от отца к сыну. Оба типа полиморфизма ДНК дополняют друг друга, давая раздельную информацию об отцовском и материнском вкладе в этническую историю и эволюцию популяций.
Передаваясь из поколения в поколение только по одной из родительских линий и не участвуя в рекомбинации (обмен участками гомологичных хромосом в процессе мейоза), оба типа полиморфизма позволяют, по крайней мере теоретически, реконструировать генетические события от наиболее популярных предков современного человека - "Y-хромосомного Адама" и "митохондриальной Евы" - до современных популяций. Полиморфизм маркеров мтДНК и Y-хромосомы определяется факторами микроэволюции (миграция, отбор, мутации). Однако характер их вариабельности по-разному отражает действие и результат этих процессов.
Уровень разнообразия геномов представителей какого-либо биологического вида зависит, во-первых, от разнообразия геномов прародителей вида и, во-вторых, от скорости накопления случайных "ошибок" (мутаций), возникающих при "переписывании" клеткой генетических текстов, и еще от того, как долго существует данный вид. При сравнении генетических текстов Y-хромосомы (или мтДНК) разных людей по присутствию в них одинаковых мутаций можно выявить общего предка. Согласно современным представлениям, скорость накопления мутаций в ДНК относительно постоянна, большинство мутаций нейтральны, так как не затрагивают значимые, смысловые участки генома. Поэтому они не отсеиваются отбором и, раз появившись, передаются из поколения в поколение. Сравнивая два родственных генетических текста, по количеству различий между ними можно установить время появления как мутаций, так и общего предка по мужской или женской линиям.
Изучение полиморфизма ДНК позволяет выявлять значительные внутри- и межпопуляционные различия в частотах полиморфных маркеров ДНК во многих географических районах мира, что стало одной из важнейших характеристик генетической структуры человеческих сообществ. За последнее десятилетие генетиками собраны и проанализированы коллекции мтДНК и Y-xpoмосом представителей народов почти всего мира. По ним восстановлена последовательность и время появления мутаций в ДНК человека.
ДНК-маркеры - эффективный инструмент для исследования гаплотипов - сочетаний аллелей тесно сцепленных полиморфных локусов. Аллель - одна из возможных альтернативных форм гена, а локус - область локализации гена в хромосоме или молекуле ДНК. Такие хромосомные участки (гаплотипы) весьма невелики по размерам, поэтому очень редко рекомбинируют. Они ведут себя как единые блоки, мало меняющиеся во времени и поэтому имеющие довольно древнее происхождение. Таким образом, размер сохранившегося неизменным гаплотипа может служить мерой времени, которое прошло от какого-то момента в прошлом. В общем случае суть анализа гаплотипов состоит в поиске сцепления между собой локусов вследствие эффекта основателя. Анализ частоты и возраста появления в популяции гаплотипа позволяет проследить его историю, а вместе с ней и события, сопутствующие его распространению. Очевидно, что такие данные представляют большой интерес для изучения истории современных народов, характеристики генофондов и оценки основных направлений эволюции всего человечества.
В 80-90-х годах прошлого века шло интенсивное накопление знаний об изменчивости мтДНК человека, были описаны основные расовые и по-пуляционно-специфические типы мтДНК. Глобальный скрининг всех основных расовых групп человечества по полиморфизму митохондриаль-ного генома позволил выявить наиболее древние мутации - ключевые для определения расоспеци-фических кластеров. Установлено, что митохонд-риальные геномы представлены комбинациями расовых групп типов мтДНК, каждая из которых ведет происхождение от единственного основателя.
Сейчас изменчивость митохондриального генома изучается на качественно новом уровне. Анализ однонуклеотидных замен мтДНК, рыяв-ляемых классическим методом анализа полиморфизма ДНК, в сочетании с изучением нуклеотид-ной последовательности гиперизменчивой области мтДНК позволяет проводить комплексную оценку полиморфизма мтДНК в популяциях человека. Такой подход выявил важную для изучения молекулярной эволюции особенность мито-хондриальной ДНК: определенным расовым группам типов мтДНК, ключевые мутации которых расположены в различных участках молекулы, соответствуют вполне определеные типы нуклеотидных последовательностей гипервариа-бельного участка мтДНК. Поскольку существует неравновесие по сцеплению между мутациями в митохондриальном геноме, молекулу ДНК можно рассматривать как один локус, представленный множеством аллелей, определенные группы которых соответствуют определенным группам сцепления между конкретными мутациями. Эти два подхода положены в основу классификации и эволюции линий всего митохондриального генома современного человечества.
ГЕНОМНЫЙ КЛЮЧ К ПРОБЛЕМЕ ПРОИСХОЖДЕНИЯ НАРОДОВ
Первым приложением ДНК-маркеров к проблеме происхождения и расселения человечества стали выполненные Канном с коллегами исследования митохондриальной ДНК представителей различных рас - африканцев, европейцев, азиатов, австралийцев и жителей Новой Гвинеи. По количеству замен нуклеотидов в мтДНК была определена степень родства различных групп людей и построено эволюционное древо человечества (рис. 1). Самая ранняя точка ветвления на древе отделяет от остальных людей группу африканцев, что указывает на африканское происхождение Homo sapiens. Именно в Южной Африке найдены самые древние мутации у бушменов и готтентотов и зарегистрировано самое высокое разнообразие мтДНК.
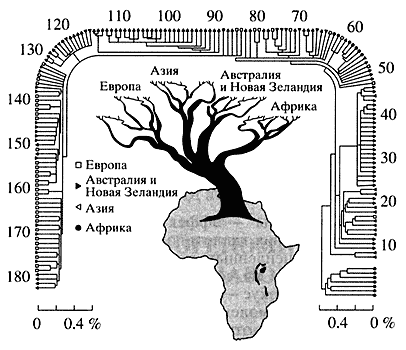
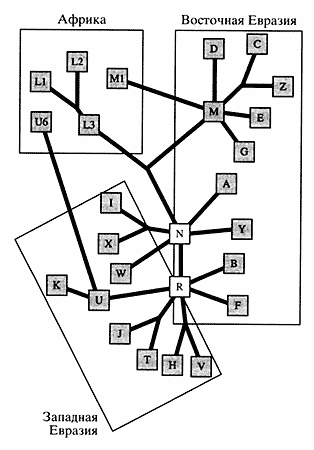
Рис. 1. Эволюционное древо человечества.
Числа соответствуют количеству исследованных популяций;
проценты - уровню внутрипопуляционного разнообразия,
оно самое высокое (0.6%) в африканских популяциях и самое низкое (0.2%) - в европейскихМитохондриальные ДНК у населения других континентов менее разнообразны, и сравнение их с мтДНК аборигенов Южной Африки показало, что они возникли как мутационные изменения африканских типов после того, как человечество распространилось за пределы этого континента. Второй вывод касался времени коалесценции (расхождения) митохондриальной ДНК. По дате отделения ветви шимпанзе (5-7 млн. лет назад), приняв темп мутационной дивергенции равным 2-4% за 1 млн. лет, Канн с коллегами вычислили время существования последней предковой мтДНК, общей для всех ныне живущих людей, - примерно 185 тыс. лет назад.
Последующие работы подтвердили африканские корни древа мтДНК современного человечества, хотя и остаются еще отдельные спорные моменты. По незавивимым оценкам нескольких групп исследователей, "митохондриальная Ева" жила в период резкого сокращения численности наших предков (до 10 тыс.), вызванного, по-видимому, изменениями климата, - 185 тыс. лет назад. Именно этот период считают временем появления Homo sapiens как биологического вида. Сравнительное исследование митохондриальной ДНК разных популяций современных людей позволило выдвинуть предположение, что еще до выхода из Африки (около 60-70 тыс. лет назад) предковая популяция разделилась по крайней мере на три группы, давшие начало трем расам - африканской, монголоидной и европеоидной [1].
Вскоре были построены генеалогические древа по данным изучения ДНК Y-хромосомы [2]. Исследование небольшого участка Y-хромосомы свидетельствует о возможно гораздо более позднем происхождении "Y-хромосомного Адама" - 140-175 тыс. лет назад. Результаты всех исследований указывают на его африканское происхождение. Различия между оценками, базирующимися на мтДНК и Y-хромосоме, могут быть объяснены как несходством демографической истории популяций по мужской и женской линиям, различным поведением женщин и мужчин при переселениях, завоеваниях и колонизациях, так и различиями самих геномов, например, в интенсивности отбора вариантов мтДНК и Y-хромосомы.
Гипотезу африканского происхождения современного человека подтверждает и наибольший уровень наследственного разнообразия в Африке по сравнению с другими континентами, а также малые различия между популяциями (на долю межпопуляционного разнообразия приходится 10-15% геномной вариабельности), что отражает недавнее происхождение биологического вида [3]. В целом массив геномных данных наиболее соответствует гипотезе недавнего африканского происхождения современного человека и доказывает справедливость монофилетической гипотезы. В то же время генетические данные не являются исчерпывающим и бесспорным доказательством этой гипотезы.
На основе распределения у разных народов частот различных мутаций в Y-хромосоме и мтДНК составлена карта расселения людей с африканской прародины [4]. Первые волны расселения человека современного типа прошли из Африки через Азию в Австралию и Европу. Удивительно, что время расселения человека по разным континентам соответствует датировке сделанных на этих континентах археологических находок. Например, появление человека в Австралии и Новой Гвинее датируется 50-60 тыс. лет назад, согласно генетическим данным. То же самое время показывает анализ изотопного состава химических элементов в археологических находках. В Центральной и Юго-Восточной Азии люди появились примерно 70 тыс. лет назад. Заселение Европы произошло позже, около 35-40 тыс. лет назад. Наиболее спорны оценки времени заселения Америки. Люди появились там гораздо позже, чем на других континентах, потому что нужно было пересечь Сибирь, добраться до Чукотки и воспользоваться тем моментом, когда уровень моря в период оледенения позволял перейти нынешний Берингов пролив. Случилось это в промежуток времени от 15 до 35 тыс. лет назад. Позже, под натиском ледника, палеолитические европейцы несколько раз отступали на юг и юго-восток, возможно, даже возвращались обратно в Африку, о чем свидетельствуют результаты исследования гаплотипов Y-хромосомы в популяциях Африки [5]. Сравнивая спектр мутаций в ДНК современных европейцев и их азиатских соседей, удалось установить, что 10-20% генов было привнесено в Европу неолитическими переселенцами с Ближнего Востока около 10 тыс. лет назад. Вместе с ними в Европе появилось земледелие.
Разные расы и народы возникли после разделения предковых популяций. Эволюция вновь образовавшихся популяционных групп шла независимо. В каждой из них накапливались свои мутации, увеличивалась генетическая дистанция между группами. Сообщества приспосабливались к климатическим и географическим условиям, типу питания. В изолированных группах независимо протекала эволюция языка и культуры.
На формирование современных народов влияли не только процессы разделения популяций, поскольку народы могут образовываться при смешении нескольких исходных сообществ с разной расовой и языковой принадлежностью. Тогда возникает генетически разнородная этническая общность, но с единым типом культуры и общим языком. В связи с этим все большую актуальность приобретает изучение генетической истории популяций отдельных регионов, расово-этнических групп, генетической родословной современных этносов.
ЭТНОГЕНОМИКА НАРОДОВ ВОЛГО-УРАЛЬСКОГО РЕГИОНА
В Волго-Уральском регионе столкнулись две волны расселения: европеоидная и монголоидная. Находясь на границе двух частей света - Европы и Азии, этот регион на протяжении исторически длительного времени был местом взаимодействия многих этнических слоев. В формировании народов края известна роль угров Западной Сибири, финнов севера Восточной Европы, индо-иранцев Ближнего Востока, тюрков Южной Сибири и Алтая, а позднее кочевых татаро-монголь-ских племен и славянских народов Центральной и Западной Европы. Вплоть до XVI в. Среднее Поволжье было подлинным "котлом", в котором перемешались многие этнические группы. Следы взаимопроникновения хранят гены проживающих здесь народов.
Современные популяции Волго-Урала крайне неоднородны по этнолингвистической структуре. Сейчас здесь проживают представители финно-угорской ветви Уральской языковой семьи (удмурты, марийцы, мордва, коми), тюркской ветви Алтайской языковой семьи (башкиры, татары, чуваши) и восточно-славянской ветви Индоевропейской языковой семьи (русские). С целью познания факторов формирования генофонда, генетической реконструкции родства и происхождения современных народов Волго-Урала изучены 17 полиморфных аутосомных ДНК-локусов, определена нуклеотидная последовательность ДНК гипервариабельного участка митохондриального генома, выполнен анализ полиморфизма 24 локусов Y-хромосомы. В ходе исследований проанализированы геномы 1500 индивидов из 8 популяций (башкиры, татары, русские, чуваши, удмурты, коми, мордва, марийцы).
Полиморфизм аутосомных ДНК-локусов. Анализ полиморфизма 17 аутосомных ДНК-локусов в популяциях Волго-Уральского региона свидетельствует об их неоднородности и высокой информативности для популяционно-генетических исследований. Оценка степени генного разнообразия (Gst) народов этого региона показала, что их генофонд не только географически занимает промежуточное место между европейскими и сибирскими генофондами. Уровень генетических различий между этносами Волго-Урала (Gst = 1.91%) занимает как бы промежуточное положение между европейскими (Gst = 1.18%) и сибирскими народами (Gst = 5.84%), тяготея к европейскому типу (рис. 2). Изучение соотношения европеоидного и монголоидного вкладов в генофонд народов Волго-Уральского региона по данным полиморфизма ДНК-локусов выявило большую долю европеоидности. Самая высокая доля европеоидности обнаружена в популяциях мордвы (89.4%) и коми (81.0%), самая низкая - в популяциях башкир (62%) и марийцев (53%) [6]. Это согласуется с данными антропологии и археологии.
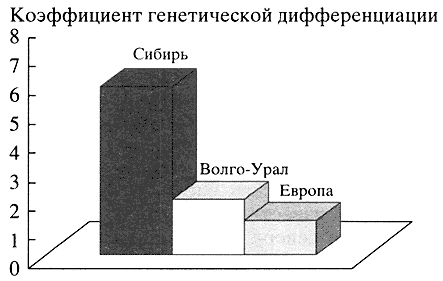
Рис. 2. Уровень генетических различий между этносами Сибири, Волго-Уральского региона и Европы
Полиморфизм мтДНК. Чтобы судить о генетическом разнообразии народов Волго-Уральского региона по материнской линии, была определена последовательность гипервариабельного участка митохондриальной ДНК (377 нуклеотидов) и проведен анализ полиморфизма мтДНК с использованием 26 эндонуклеаз (ферментов рестрикции). Полученные данные анализировались методом так называемых медианных сетей. Медианная сеть - один из графических вариантов представления данных о полиморфизме ДНК, который позволяет проследить дивергенцию митохондриального генома от предковой ДНК. В качестве предкового выбирается гаплотип, который широко распространен в данной популяции и включает наиболее частые аллели, характерные для данного этноса. В процессе такого анализа в единую медианную сеть соединяются посредством линий сотни и тысячи соседствующих, наиболее близких индивидуумов. В этой сети узловые соединения совпадают со специфическими гаплотипами, определенными по характерным мутациям изучаемого генома.
При сравнительном изучении мтДНК у народов Волго-Уральского региона выявленные гап-лотипы были отнесены к определенным линиям в соответствии с классификацией Макуолли [7]. Для большинства расовых и географических групп установлена специфичность типов мтДНК (рис. 3). Более 90% всех типов мтДНК народов Западной Евразии принадлежит к линиям Н, J, V, 1, К, Т, U, W и X, которые рассматриваются как потомки европейского генного пула, существовавшего в верхнем палеолите. Митохондриальный геном азиатских популяций включает два суперкластера М и N, которые соединяются в африканском макрокластере L3. Последний рассматривается в качестве общего предка для всех неафриканских популяций мира. Около 50% линий мтДНК коренных жителей Азии принадлежит к суперкластеру М, который подразделяется на линии С, Z, D, G, Е и др. [8]. Суперкластер N в Азии делится на несколько линий: А, Y, В, F.
Рис. 3. Медианная сеть, демонстрирующая полиморфизм мтДНК в популяциях Европы, Азии и АфрикиЛинии D, С, Z, G суперкластера М, линии A, Y суперкластера N и линии В, F суперкластера R характерны для азиатских популяций; линии 1, X, W суперкластера N и линии V, Н. Т, J, U, К суперкластера R - для европейских; линии LI, L2, L3, М1 и U6 принадлежат африканским народам
Большинство типов мтДНК народов Волго-Уральского региона соответствует линиям мтДНК Европы и Ближнего Востока, что свидетельствует об общих предковых линиях мтДНК, специфичных для европейцев [9]. В целом среди изученных нами популяций частота европейских типов мтДНК оказалась наиболее высокой у мордвы, коми-зырян и русских. С другой стороны, уровень распространения линий мтДНК, специфичных для Восточной Евразии, также достигает больших значений, что ранее не было показано для Западной Европы. Высокая частота линий G, D, С, Z и F в некоторых этнических группах, как тюркских (башкиры), так и финноугорских (удмурты, коми-пермяки), указывает на значительное участие сибирского и центральноазиатского компонента в этногенезе народов Волго-Уральского региона.
Самостоятельный интерес вызывает высокая частота азиатской линии F (6%) у башкир. Эта линия характерна для народов Средней Азии - казахов, уйгуров и монголов, и мы можем предположить, что, во-первых, существенную роль в формировании данной этнической группы сыграл среднеазиатский компонент и, во-вторых, популяция башкир длительное время находится в изоляции от своих ближайших соседей. У других тюркоязычных и финноугорских популяций Волго-Уральского региона частота азиатских линий низкая. Поскольку очевидных географических барьеров как между тюркскими этносами, так и между финноугорскими популяциями не существует, можно сделать вывод о различной демографической истории башкир по материнской линии внутри упомянутых языковых семей. Выборки башкир и удмуртов по совокупности всех полученных по материнской линии данных можно охарактеризовать как этносы, имеющие в прошлом период резкого роста численности в условиях относительной изоляции. Анализ митохондриального генома татар, чувашей, марийцев, мордвы, коми и русских, скорее всего, отражает процессы продолжающейся интенсивной метисации в условиях сохранения постоянного популяционного размера. В целом, медианные сети показывают смешение и взаимопроникновение гаплотипов мтДНК, что свидетельствует как о тесных этногенетических контактах изученных этносов, так и о единой генетической основе населения Волго-Уральского региона по материнской линии.
В распространении типов мтДНК у народов Волго-Уральского региона ведущую роль играют факторы этнокультурной и территориальной близости или отдаленности, но не лингвистические барьеры. Это означает, что по материнской линии финноугорские народы имеют большее сходство с их непосредственными тюркскими соседями, чем с родственными в языковом отношении балтофинскими народами.
Анализ европеоидного и монголоидного вклада в материнские генетические линии народов Волго-Уральского региона не выявил корреляции языка и геномного состава этнических групп. На языках тюркской группы, привнесенных из Азии, говорят не только башкиры (65% монголоидности), но и татары и чуваши, у которых преобладает европеоидный генетический компонент. В остальных популяциях региона вклад монголоидного компонента составляет от 12% у русских до 20% у удмуртов. Русские, проживающие на территории этого региона, имеют 10-12% монголоидных типов мтДНК, а русские из Рязанской и Курской областей - только 2-3%. Это можно объяснить смешением русских с тюркоязычными народами на территории Волго-Уральского региона.
Интересно, что некоторые материнские линии у разных народов, например, у русских, татар и марийцев, оказались общими. Это показывает глубокое родство народов, говорящих на разных языках, придерживающихся разных религий и традиций.
Сравнительный анализ типов мтДНК в 18 популяциях Евразии, включая популяции Волго-Уральского региона (гагаузов, турков, татар, башкир, чувашей, карачаевцев, кумыков, азербайджанцев, узбеков, казахов, киргизов, ногайцев, уйгуров, шорцев, тувинцев, долган, якутов), которые относятся к тюркской ветви Алтайской языковой семьи, позволил установить западно-восточный градиент увеличения частоты азиатских линий мтДНК на расстоянии 8000 км: от 1% у гагаузов из Молдавии до 95% у якутов и 99% у долган (рис. 4). Кроме того, установлено, что лингвистическое сходство популяций играет меньшую роль, чем географическая близость или отдаленность популяций.
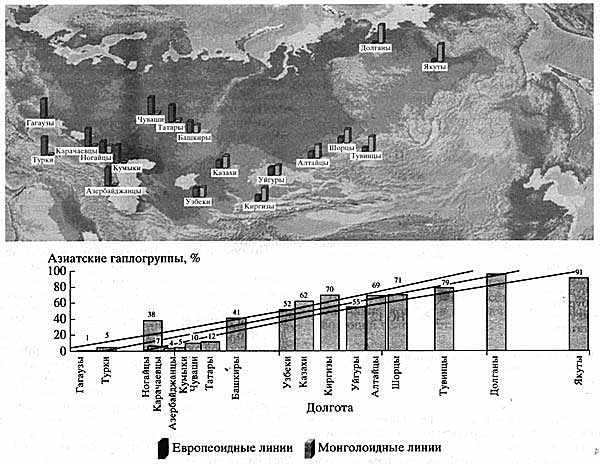
Рис. 4. Результаты сравнительного анализа типов мтДНК в 18 популяциях Евразии
Отчетливо виден западно-восточный градиент увеличения частоты азиатских линий мтДНКОдин из наиболее важных аспектов анализа митохондриального генома - оценка времени коалесценции (расхождения, дивергенции) линий мтДНК в пределах каждой линии. Безусловно, на временные оценки будут влиять различные факторы формирования разнообразия мтДНК: объем выборки, миграция населения, резкий рост численности, феномен "бутылочного горлышка" - сильное сокращение численности наших предков, вызванное, по-видимому, изменением климата, и т.д. Тем не менее оценка времени дивергенции линии возможна при обнаружении предковых гаплотипов.
По ориентировочным оценкам, возраст дивергенции линий, выявленных у народов Волго-Уральского региона, варьировал от 273 ± 57 тыс. лет для азиатской линии Z до 22.76 ± 5.250 тыс. лет для линии С. Возраст дивергенции самой крупной европейской линии Н определен в 20.036 ± 4.250 тыс. лет, что соответствует археологическому времени повторной экспансии населения на территории Урала в постледниковый период. Используя данные о числе мутационных замен и скорости накопления мутации для гипервариабельного участка мтДНК, равной одной мутационной замене за 20.18 тыс. лет, мы получили среднее значение времени дивергенеции мтДНК для народов Волго-Уральского региона. Оно составляет 49.60 тыс. лет назад, что соответствует периоду расселения человека на европейском континенте в эпоху верхнего палеолита.
Полиморфизм ДНК Y-хромосомы. Анализ Y-хромосомы вошел в арсенал методов эволюционной генетики лишь в самое последнее время, когда был найден ряд высокоинформативных полиморфных локусов в ее нерекомбинантной части. Генетические свойства Y-хромосомы, такие как передача только по отцовской линии, отсутствие рекомбинации, малая эффективная численность пула Y-хромосом по сравнению с аутосомами (в четыре раза меньше, чем у аутосом), позволяют прослеживать по гаплотипам Y-хромосомы отцовские линии, представляющие собой последовательную "запись" мутаций в ряду поколений. По сравнению с митохондриальным геномом, насчитывающим 16.5 тыс. пар нуклеотидов. Y-хромосома, размер которой оценивается приблизительно в 60 млн. пар нуклеотидов, дает в руки исследователей потенциально более мощное "оружие".
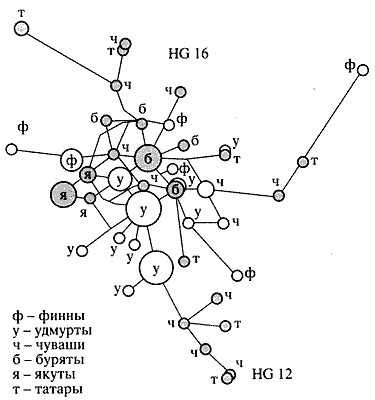
Если предшествующие работы, посвященные анализу Y-хромосомы в популяциях России, основывались преимущественно на анализе 9 маркеров, то для изучения и сравнения генетического разнообразия отцовских линий в популяциях Волго-Уральского региона использовались 24 маркера Y-хромосомы. В качестве примера на рисунке 5 представлена медианная сеть линий 12 и 16 Y-хромосомы как наиболее интересных в контексте финноугорских народов. Линия 16 практически отсутствует в западноевропейских популяциях, но частота ее высокая среди народов Балтии - эстонцев и финнов, а также у народов Волго-Уральского региона, особенно удмуртов и коми-зырян [10].
Рис. 5. Медианная сеть линий HG12 и HG16 Y-хромосомы, построенная для некоторых популяций Европы и Азии Для линии 16 Y-хромосомы уровень генетического разнообразия намного выше в популяциях Восточной Европы (чуваши, татары), чем в изученных популяциях Сибири. Хотя удмурты и имеют очень высокую частоту линий 12 и 16, уровень генетического разнообразия у них невысокий по сравнению с другими популяциями Европы. Данные о низком уровне генетического разнообразия удмуртов получены и по материнской линии при анализе полиморфизма митохондриальной ДНК. Все это свидетельствует о несомненной роли эффекта основателя и дрейфа генов в демографической истории удмуртов.
Анализ распространения и разнообразия линии 16 Y-хромосомы среди восточноевропейских популяций показывает, что местом ее "рождения", возможно, является Восточно-Европейская равнина. Согласно филогеографическому анализу этой линии в Евразии, она начала распространяться с запада на восток. В то же время частота распространения линии 12 - предковой для линии 16 - меньше, чем для линии 16.
Особенно характерна для популяций Волго-Уральского региона линия 3 Y-хромосомы, частота встречаемости которой максимальна у славян (русские и поляки), а также у населения Латвии, Литвы и Эстонии [10]. Таким образом, носители этой линии - этнические группы, принадлежащие к разным языковым семьям. Частота встречаемости таких групп уменьшается в направлении север (Финляндия, Швеция) - юг (Турция, Кавказ). Анализ распространения данной линии в популяциях Волго-Уральского региона подтверждает гипотезу о возможном движении населения после Ледникового периода (Last Glacial Maximum) с территории нынешней Украины, где в то время располагался один из центров потепления [1].
Судя по результатам анализа отцовских линий в популяциях Восточной Европы и, в частности, Волго-Уральского региона, главную роль в формировании генетического разнообразия народов, проживающих на этой территории, по-видимому, играет географическая близость, а не языковая принадлежность. И хотя многие особенности генетической близости популяций объясняются с точки зрения их географического положения, в некоторых случаях "индивидуальная" демографическая история популяции имеет существенное значение. Хороший пример - популяция удмуртов, у которых разнообразие линий Y-хромосомы и мтДНК ограничено. Принимая темпы мутирования изученных ДНК-маркеров Y-хромосомы за 2.1 х 10-3 и длительность одного поколения за 25 лет, получаем, что наблюдаемая дисперсия гаплотипов, выявленных у современного населения Волго-Уральского региона, сформировалась приблизительно 42.5 тыс. лет назад, что соответствует времени заселения Европы человеком в эпоху верхнего палеолита.
Таким образом, исследования полиморфизма аутосомных, митохондриальных и Y-хромосомных ДНК-маркеров внесли важный вклад в понимание путей происхождения человека и рас, расселения Homo sapiens по планете, в генетическую и демографическую историю отдельных этносов и популяций. Можно надеяться, что по мере все более подробного изучения свойств конкретных маркеров ДНК будут появляться дополнительные возможности для изучения генетической истории народов Европы и Азии. Дальнейшее развитие этногеномики в сочетании с палео- и археогеномикой значительно расширит наши представления о генофонде человека, внесет весомый вклад в понимание вопросов исторического развития и эволюции человечества.
ЛИТЕРАТУРА
1. Rosser Z.H., Zerjal Т., Hurles M.E. et al. Y-chromosomal diversity in Europe is clinal and influenced primarily by geography, rather than by language // Am. J. Hum. Genet. 2000. V. 67. P. 1526-1543.
2. Thomson R., Pritchard J., Shen P., Oefner P., Feldman W. Recent common ancestry of human Y chromosomes: Evidence from DNA sequence data // Proceedings of National Academy of Sciences. 2000. V. 97. №13 P. 7360-7365.
3. Stoneking M. Progress in population genetics and human evolution//Berlin: Springer, 1997. P. 164.
4. Cavalh-Sforza L.L. Genes, Peoples, and Languages. N.Y.: North Point Press, 2000.
5. Cruciani F., Santolamazza P.. Shen P. et al. A back migration from Asia to Sub-Saharan Africa is Supported by high- resolution analysis of Human Y-chromosome hap-lotypes //Am. J. Hum. Gen. 2002. V. 70. P. 1197-1214.
6. Лимборская C.A., Хуснутдинова Э.К.. Балановская Е.В. Этногеномика и геногеография народов Восточной Европы. M.: Наука, 2002.
7. Macualy V.A., Richards M.B., Forster P. et al. The Emerging Tree of West Eurasian mtDNAs: A Synthesis of Control Region Sequences and RFLPS //Am. J. Hum. Genet. 1999. V. 64. P. 232-249.
8. Wallace D.C., Brown M.D., Lott M.T. Mitochondrial DNA variation in human evolution and disease // Gene. 1999. V. 238. P. 211-230.
9. Бермишева M., Тамбетс К., Виллемс P., Хуснутдинова Э. Разнообразие гаплогрупп митохондриальной ДНК у народов Волго-Уральского региона // Молекулярная биология. 2002. № 6. С. 990-1001.
10. Villems R., Rootsi S., Khusnutdinova E. el al. Archaeo-genetics of Finno-Ugric speaking populations // The Roots of Peoples and Languages of Northern Eurasia. IV. Ed. by K. Julku. Oulu. 2002. P. 271-284.