 |
БАБОЧКИ:
|
|
БАБОЧКИ:
|
ВЗАИМОСВЯЗЬ живого организма с окружающей средой обычно изучают, исходя из биологических концепций. Однако и биологические объекты, и машины подчиняются одним и тем же физическим законам, и поэтому живое можно рассматривать с инженерной точки зрения. Жизненно важные функции организма осуществляются посредством множества различных физических механизмов. Конкретный характер этих механизмов обусловливает тот диапазон внешних условий, в которых организм может существовать и размножаться. Вызывая изменения в работе того или иного механизма, можно установить, как он влияет на распространение и эволюционный успех данного вида организмов. Я применил этот подход к весьма странным с инженерной точки зрения конструкциям - бабочкам.
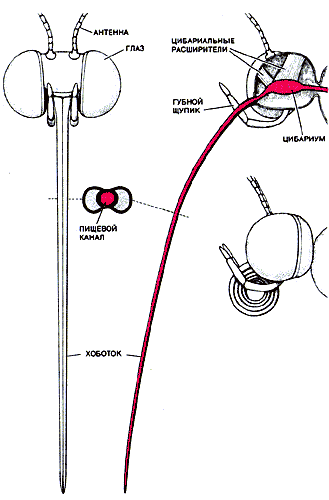
Следует сказать, что есть принципиальная разница между созданием технического описания машины и инженерным анализом живого существа. Конструктор машины исходно знает, какие функции машина должна выполнять, и разрабатывает устройство, способное осуществлять их с максимальной эффективностью или производительностью. Зоолог, избравший в своей работе инженерный подход, поступает наоборот, делая выводы о функции объекта исследования на основании его анатомического строения. В отличие от машин живые организмы обладают способностью самопостроения, и все новые структуры представляют собой видоизменения уже существующих. Встав на позиции инженера, биолог должен изучать ту или иную структуру не отдельно взятую, а обязательно с учетом эволюционных отношений между организмами и тех процессов развития, в результате которых данная структура возникла. С помощью инженерных методов анализа можно подойти к пониманию обоих этих типов биологического взаимодействия. Три наиболее важные функции, обеспечивающие выживание и размножение бабочек, - это питание, теплорегуляция и полет. Полет, играющий центральную роль в биологии бабочек, является энергетически весьма дорогостоящим способом передвижения, требующим много высокоэффективного топлива. Источником топлива служит нектар, которым питается бабочка. Нектар образуется в цветках многих растений и представляет собой смесь воды с простыми сахарами. В то время как у большинства насекомых ротовой аппарат состоит из нескольких отдельных частей, у бабочек эти части слились в единую структуру, именуемую хоботком. Хоботок имеет вид длинной гибкой трубки: когда бабочка находится в состоянии покоя, он плотно свернут, а во время питания развертывается и его кончик погружается в нектар.
В голове бабочки имеется полость, размер которой регулируется специальными мышцами, носящими название цибариальных расширителей. Сокращение этих мышц создает давление, заставляющее нектар подниматься по длинному тонкому хоботку. С инженерной точки зрения способ питания бабочки аналогичен всасыванию раствора сахара через длинную соломинку. Всякому любителю коктейлей известны по крайней мере два фактора, влияющие на питье через соломинку. Во-первых, чем длиннее и тоньше соломинка, тем больше должно быть давление, чтобы заставить жидкость подниматься по ней. Во-вторых, это давление должно увеличиваться с повышением вязкости жидкости.
Каким образом бабочка добивается быстрого поступления энергии в свой организм? Содержание сахара в нектаре колеблется очень сильно, и можно было бы ожидать, что быстрее всего насекомое получит энергию, высасывая наиболее сладкий из доступных ему нектаров. Но это не так. Действительно, в процессе всасывания нектара с низким содержанием сахара поглощение энергии будет происходить медленно. Однако и очень сладкий нектар не обеспечивает максимально быстрого поступления энергии. Причина здесь в том, что с повышением концентрации сахара быстро увеличивается вязкость раствора. Если нектар очень сладкий, он не может засасываться достаточно быстро, чтобы насекомое получало энергию с максимально высокой скоростью. Поэтому для бабочки оптимален такой нектар, который содержит не наибольшее, но и не наименьшее количество сахара.
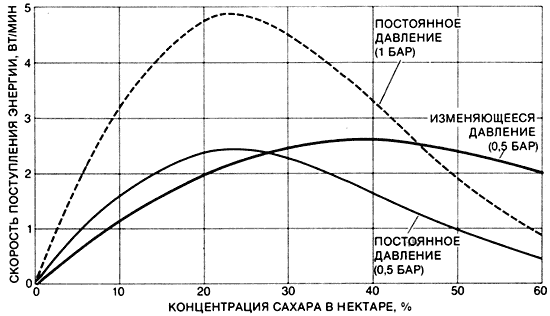
Оптимальная концентрация сахара в нектаре, которым питается бабочка, зависит от механизма, обеспечивающего его всасывание. Оптимальнойявляется концентрация, которая позволяет бабочке максимально быстро получать необходимую энергию. На графике показано, как изменяется скорость поступления энергии в зависимости от концентрации сахара для трех несколько различающихся механизмов высасывания нектара. Если в хоботке создается низкое постоянное давление, оптимальна концентрация 20-25% (сплошная черная пиния). Если давление постоянное и сравнительно высокое, то скорость поступления энергии при всех концентрациях больше, но оптимальная величина остается той же (пунктирная черная пиния). Если же давление всасывания растет с увеличением концентрации, оптимальное содержание сахара в нектаре составляет примерно 35% (цветная линия). Кривые для постоянного давления являются гипотетическими.Кривая для изменяющегося давления построена по данным П. Мэя из Университета Флориды для бабочки-геликониды Agraulis vanillae. Самый сладкий нектар вовсе не наилучший для бабочек, поскольку с увеличением концентрации сахара увеличивается вязкость нектара, и его становится труднее сосать.Какое содержание сахара в нектаре будет оптимальным, зависит от особенностей механизма, обеспечивающего процесс питания бабочки. Т. Дэниел из Вашингтонского университета, П. Мэй из Университета Флориды и я смоделировали два механизма: в одном из них давление всасывания, создаваемое бабочкой, постоянно и независимо от содержания сахара в нектаре, а в другом - давление изменяется с изменением концентрации сахара. Оказалось, что если давление сохраняется постоянным, то оптимальным будет нектар, содержащий 20-25% сахара, и при этом несущественны размеры и форма хоботка. Увеличение диаметра хоботка или уменьшение его длины увеличивает скорость засасывания нектара, но так как это имеет место при всех концентрациях сахара, то оптимальная концентрация остается неизменной.Таким же образом, если бы бабочка могла вдруг увеличить вдвое давление всасывания при всех концентрациях сахара, скорость поступления энергии при всасывании, скажем, 40%-ного раствора резко увеличилась бы, но она увеличилась бы пропорционально и в случае 25%-ного раствора. Напротив, если давление удвоить для 25%-ного раствора и утроить для 40%-ного, то во втором случае бабочка будет получать энергию быстрее. И действительно, наши исследования показали, что когда давление всасывания изменяется в зависимости от концентрации сахара в нектаре, оптимален нектар с концентрацией сахара от 35 до 40%. Какой из этих двух механизмов действует, у большинства бабочек неизвестно. По-видимому, единственные опубликованные данные - это результаты П. Мэя, который показал для двух видов бабочек, что они пользуются механизмом с изменением давления всасывания в зависимости от концентрации сахара в нектаре. Вполне вероятно, что быстрота получения "прибыли" может быть важным фактором в выборе бабочкой цветов, которые она посещает. Бабочки стараются питаться на растениях, дающих нектар с содержанием сахара от 15 до 30%. Пчелы в отличие от бабочек посещают растения, цветки которых образуют нектар с концентрацией сахара 10-50%.
Даже небольшое изменение в физическом механизме, обеспечивающем процесс питания, очень сильно влияет на отношения между организмом бабочки и окружающей средой. То же самое можно сказать и о теплорегуляции. Для того чтобы летать, многие бабочки должны поднимать температуру тела до 30-40°С, т.е. до величины, обычной для большинства млекопитающих. Млекопитающие согревают себя путем увеличения интенсивности метаболизма, которое ведет к усилению выделения тепла, а "охлаждаются", регулируя приток крови к поверхности тела. У бабочек механизмы терморегуляции совершенно иные. В теле насекомых также имеется внутренняя жидкость - гемолимфа, - но ее ток не регулируется столь тонко. И чтобы поднять свою температуру, бабочка не увеличивает интенсивность обмена веществ, а просто подставляет тело солнечным лучам, поглощая солнечную энергию до тех пор, пока не сможет подняться в воздух.
Экспозиция с поглощением: крылья бабочки поглощают солнечные лучи и передают тепловую энергию телу. Для такого способа согревания существуют две основные позы. При экспозиции боком (латеральная поза) бабочка держит сложенные вместе крылья над телом {слева). При экспозиции спинкой (дорсальная поза) крылья раскрыты и направлены в стороны (справа). Каждому виду, как правило, свойственна какая-то одна из этих двух поз. В зависимости от того, какую позу принимает бабочка, для температуры тела имеет значение цвет либо верхней, либо нижней поверхности крыльев. Бабочки из рода Colias, изображенные следующем рисунке, принимают при экспозиции латеральную позу.Известно несколько поз, которые принимают бабочки, греясь на солнце. Для каждого вида обычно характерна одна определенная поза. Эта поза зависит от специфики того механизма, который обеспечивает нагревание тела. Когда бабочка сидит под лучами солнца, тепловая энергия сообщается телу одним из двух способов: либо крылья поглощают солнечное излучение и тело бабочки нагревается посредством механизма теплопроводности (через ткани крыльев), либо крылья отражают солнечные лучи так, что они попадают на поверхность тела и поглощаются.Виды, использующие поглощение солнечных лучей крыльями, во время экспозиции принимают дорсальную или латеральную позу. В дорсальной позе крылья расправлены в стороны от тела, как у самолета. В латеральной позе крылья сложены и располагаются вертикально по отношению к телу. Независимо от того, какую из этих поз выбирает бабочка, при таком способе согревания повышение температуры тела определяется количеством лучей, попадающих на ту часть крыла, которая ближе всего к телу, так как неплотные и тонкие крылья бабочки являются плохими проводниками тепла.
Так как от цвета поверхности зависит длина волны поглощаемого света, у бабочек окраске крыльев принадлежит важная роль в терморегуляции. Цвет крыльев у них создается пигментацией не самого крыла, а множества тонких чешуек, покрывающих его наподобие черепицы (см. Nijhout Н. F. The Color Patterns of Butterflies and Moths, "Scientific American", November, 1981). Каждая чешуйка содержит только один пигмент, так что в целом окраска крыла бабочки складывается для восприятия по принципу пуантилизма, как в живописи Жоржа Сера.
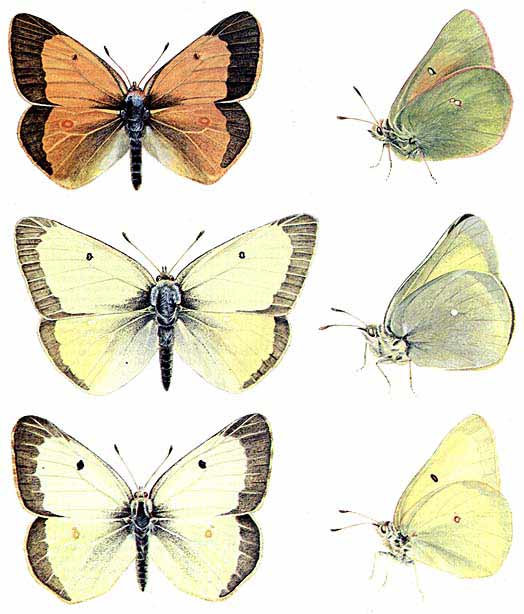
Три вида бабочек из рода Colias имеют разную окраску крыльев в зависимости от высоты, на которой они обитают в Скалистых горах. Вверху - бабочка из популяции вида Colias meadii, встречающейся на высотах 3300-3600 м. В среднем ряду - бабочка из популяции вида Соlias alexandra, который находят на высотах 2700-2900 м. Внизу - бабочка из популяции Colias philodice eriphyle, обитающей на высотах 1500 - 1700 м. Бабочки рода Colias согревают тело до температуры, необходимой для полета, располагаясь на солнце так, что крылья сложены вместе над телом, и энергия солнечных лучей, поглощенных нижней поверхностью крыльев, посредством механизма теплопроводности передается телу. Область крыла, ближайшая к телу, наиболее важна для повышения его температуры, и она тем темнее, чем на большей высоте обитает бабочка.Поскольку наибольшее значение для повышения температуры организма насекомого имеет та часть крыла, которая непосредственно примыкает к телу, окраска этой части важнее для терморегуляции, чем окраска других участков крыла. У большинства бабочек с латеральным или дорсальным типом экспозиции основание крыла имеет черный цвет благодаря присутствию пигмента меланина. Глубина оттенка зависит от количества пигмента. У. Уотт из Станфордского университета и я изучали окраску крыльев у нескольких видов бабочек рода Colias, которых часто называют желтушками. Мы установили, что у желтушек, обитающих на возвышенностях, основания крыльев темнее, чем у бабочек, живущих в более низкой местности. Темные крылья лучше поглощают солнечные лучи, что позволяет бабочкам, обитающим на большей высоте, т.е. в более холодных местах, полнее использовать солнечную энергию.Совершенно иная техническая задача решается при утилизации энергии солнечных лучей путем их отражения от поверхности крыла на тело бабочки. Поглощение и отражение света связаны обратной зависимостью. Темные поверхности хорошо поглощают свет, но плохо его отражают.
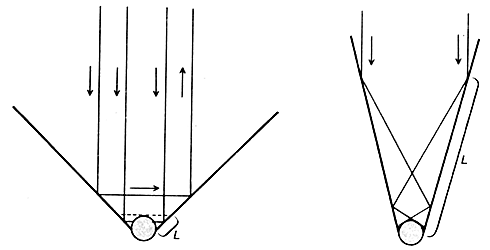
Экспозиция с отражением: солнечные лучи отражаются от верхней поверхности крыльев, попадают на тело бабочки и поглощаются. Крылья располагаются над телом буквой V. Угол между крыльями может изменяться в широких пределах. У некоторых видов он велик (слева). В этом случае только те лучи, которые падают на небольшой участок поверхности крыла у самого его основания (L), отражаются на тело бабочки. У других видов крылья образуют более узкий угол (справа) и площадь, с которой лучи попадают на тело, больше. Если угол очень мал, то на тело попадают даже лучи, отраженные вершиной крыла.Чтобы эффективно отражать солнечные лучи крылья должны быть белыми или серебристыми. И действительно, основная группа бабочек, согревающихся на солнце с помощью отражения лучей света от поверхности крыльев, даже называется белянками из-за окраски верхней поверхности крыльев (к белянкам, например, относится род Pieris.) Бабочки этой группы обычно держат крылья под углом к телу, наподобие буквы V. Величина угла между крыльями существенно влияет на эффективность попадания на тело солнечных лучей, отраженных от крыльев.Солнечные лучи, падающие на внутреннюю поверхность раздвинутых буквой V крыльев, отражаются и часть их попадает на тело бабочки, а часть уходит в сторону, не коснувшись тела. Тела достигают лишь те лучи, которые падают на ближайшую к нему часть крыла. Площадь этого участка определяется углом между крыльями. Если угол велик, только лучи, отраженные от небольшого участка у самого основания крыла, коснутся поверхности тела. Если угол маленький, на тело попадают лучи, отраженные от большего участка поверхности крыльев, иногда включая даже и вершину крыла. У каждого вида рода Pieris есть своя характерная поза для согревания под солнечными лучами, и этой позе соответствует окраска крыльев: существует некоторая связь между оттенком вершины крыла и той ролью, которую она играет в регуляции температуры тела.
Способность повышать температуру тела до величины, необходимой для осуществления полета, занимает центральное место в биологии многих видов бабочек. Для проявления всех видов жизнедеятельности, включая поиски пищи, партнера для спаривания и мест для откладки яиц, нужно, чтобы бабочка могла летать. В ряде исследований последних лет, проведенных в США и Европе, было показано, что успех размножения помимо всего прочего ограничивается временем, в течение которого бабочка может летать в поисках растений, подходящих для откладки яиц. Одна из причин этого - краткость жизни взрослой особи; у многих видов продолжительность взрослой стадии составляет менее недели. Сможет ли бабочка повысить температуру своего тела настолько, чтобы полет стал возможен, зависит более всего от погодных условий, которые, таким образом, оказывают серьезное влияние на эффективность репродуктивного процесса бабочек.
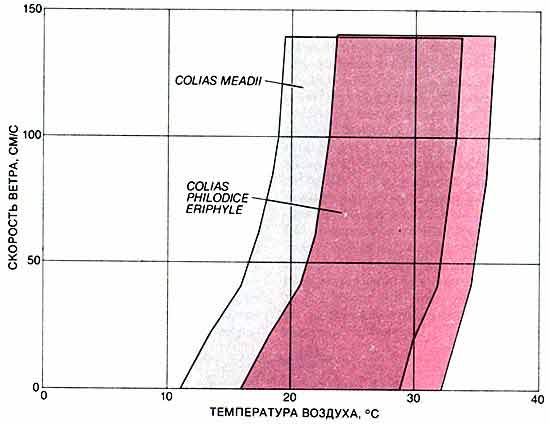
Стремясь использовать преимущества инженерного подхода, Р. Моффет из Станфордского университета и я построили математические модели, отражающие связи между физическими свойствами, влияющими на терморегуляцию (например, окраской крыльев) и погодными условиями, при которых данный вид может летать. Эти связи могут быть изображены в графической форме, предложенной У. Портером из Висконсинского университета в Мадисоне и Д. Гейтсом из Мичиганского университета, в виде "летного пространства" - области в системе координат температура воздуха-скорость ветра, охватывающей все условия, при которых бабочка данного вида может набрать достаточное для взлета количество тепловой энергии.
Летное пространство представляет собой изображенный графически диапазон климатических условий, в которых данный вид бабочек может достигать температуры тела, необходимой для полета. Специфическая окраска крыльев имеет адаптивное значение. Чтобы убедиться в этом, сравним летные пространства двух видов. Серым цветом показано летное пространство Colias meadii, обитающих на больших высотах (на одном из верхних рисунков этот вид изображен вверху). Темная нижняя поверхность крыльев позволяет этому виду летать в условиях сравнительно холодного климата. Цветное поле - летное пространство Colias philodice eriphyle, обитающих на меньших высотах (на одном из верхних рисунков этот вид изображен внизу). Светлая окраска нижней поверхности крыльев вполне соответствует более теплым климатическим условиям, в которых живут эти бабочки.При сравнении летных пространств различных видов можно видеть, как окраска крыльев связана с диапазоном климатических условий, при которых бабочка может подняться в воздух. Сравним, например, летные пространства двух видов из рода Colias, изучением которых занимались мы с Уоттом. Некоторые популяции Colias philodice eriphyle обитают на высотах 1500-1800 м над уровнем моря. Популяции же Colias meadii, у которых основания крыльев более темные, встречаются на высотах 3300-3600 м. Изобразив их летные пространства, мы увидим, что одним из средств приспособления к конкретным внешним условиям у этих близких видов является окраска крыльев. Кроме того, математическая модель дает возможность предсказать, что бабочки вида, встречающегося на меньших высотах, не смогли бы летать, попав в условия высокогорья, поскольку им не удалось бы поднять температуру своего тела до уровня, необходимого для полета. Результаты экспериментов, в которых я переносил особей С. рeriphyle на высокие горные склоны, где холодно, подтвердили это предсказание.
Как выяснилось, некоторые механизмы, обеспечивающие полет крупных бабочек, совершенно отличны от тех, которые используются в летательных аппаратах. Только недавно в результате лабораторных исследований мы начали постигать физические принципы полета бабочек. Многие крупные бабочки летают, порхая и планируя, причем периоды порхания чередуются с периодами планирования. Сочетание порхания с планированием не часто встречается у насекомых, хотя в той или иной степени этот способ используют некоторые прямокрылые и стрекозы.
Механика планирующей фазы полета бабочек изучена лучше, чем фазы порхания. Планирование понятнее нам отчасти оттого, что здесь легко воспользоваться результатами классической аэродинамики, в частности аэродинамики профиля крыла. При планирующем полете крылья фиксированы в определенном положении. Сочетание формы крыла и угла атаки относительно потока воздуха таково, что над верхней поверхностью крыла воздух движется с более высокой скоростью, чем под нижней поверхностью. Из-за разницы в скорости движения воздуха у верхней и нижней поверхностей крыла получается разница в давлении на эти поверхности, что в свою очередь создает силу, направленную вверх и перпендикулярно к направлению движения аппарата. Эта сила называется подъемной.
Структура течения около крыла во время планирующего полета весьма сложна. Чтобы в ней разобраться, полезно представить поток воздуха в виде двух составляющих: поступательной и циркуляционной. Поступательная составляюшая появляется согласно принципу обращения движения: крыло считается неподвижным, а воздух - натекающим на него. Циркуляционное течение - это круговое движение воздуха вокруг крыла. Как мы увидим дальше, прием разложения потока воздуха на составляющие, несмотря на некоторую его условность, оказывается очень полезным для понимания механизма одной из форм полета бабочек - зависания.
При планирующем полете крылья бабочки создают подъемную силу в значительной мере так же, как крылья самолета. Однако между самолетом и живым организмом имеется и существенная разница. Основная сила, препятствующая движению крыла вперед, - это сила сопротивления. Она складывается из силы трения поверхности крыла о воздух и результирующего перепада давлений, действующих на наветренную и подветренную поверхности крыла. Эффективность аэродинамического профиля крыла измеряется отношением подъемной силы к силе сопротивления. Конструкторы летательных аппаратов стараются добиться максимальной величины этого отношения. Крыло бабочки характеризуется низкой величиной отношения подъемной силы к силе сопротивления. Однако важной характеристикой планера является время, в течение которого он может оставаться в воздухе. По такому критерию бабочку следует считать хорошим планером. Когда бабочка планирует, она снижается медленно. Скорость снижения мала благодаря особой форме ее крыла, которое можно сравнить с парашютом. Сводчатая форма парашюта и большая площадь поверхности, перпендикулярной направлению снижения, создают большую силу сопротивления, противодействующую силе земного притяжения. Точно так же действуют широкие плоские крылья бабочки. Собственно, планирующий полет бабочек правильнее было бы назвать парашютированием. Когда бабочка перемещается в воздухе, то большая сила сопротивления оказывается одновременно причиной как низкой величины отношения подъемной силы к силе сопротивления, так и низкой скорости снижения.
Фаза порхания сложнее, чем фаза планирования. Разобраться в ней поможет анализ такой формы порхания, как зависание. При зависании нет чисто горизонтального или чисто вертикального перемещения бабочки в целом, хотя и крылья, и тело меняют свое положение. Бабочки, и дневные и ночные, относятся к большой группе животных (включающей многих других насекомых, а также колибри), способных порхать с зависанием. При зависании движение крыльев составляет сложный цикл, который многократно повторяется, благодаря чему птица или насекомое держится в воздухе.
Если бы можно было замедлить эти быстрые движения и наблюдать их со стороны, то мы бы увидели, что при зависании у большинства животных (в том числе у колибри и многих насекомых) крыло двигается в основном назад и вперед, а амплитуда его движения по вертикали мала. В таком случае говорят о движении с горизонтальным взмахом крыла. Рассмотрим цикл с горизонтальным взмахом применительно к профилю крыла. Профиль при движении описывает восьмерку, длинная ось которой лежит в горизонтальной плоскости. Суммировав мгновенные значения подъемной силы за весь цикл и разделив на время его осуществления, мы получим среднюю подъемную силу, которая позволяет животному держаться в воздухе.
У бабочек же движения крыльев при зависании совсем иные. Ч. Эллингтон из Кембриджского университета недавно показал, что крылья бабочки при этом перемещаются по вертикали с большой амплитудой, т.е. имеет место движение с вертикальным, а не с горизонтальным взмахом крыла. Хотя крыло бабочки двигается в основном в вертикальной плоскости, в нем есть сходство со взмахами лодочного весла. При махе вниз крыло располагается перпендикулярно направлению своего движения, когда же оно поднимается вверх, то бабочка держит его параллельно направлению движения. Бабочки единственные из известных животных используют вертикальный взмах при зависании.
Мы уже условились выше, что течение около крыла можно представить в виде двух составляющих. Однако традиционные методы анализа становятся неправомерными в случае движения с вертикальным взмахом. Чтобы вычислить суммарную величину подъемной силы, создаваемой аэродинамическим профилем крыла за весь цикл (подобно тому, как это можно сделать для взмахов в горизонтальной плоскости), необходимо допустить, что в каждый данный момент времени силы, действующие на крыло, не зависят от сил, действовавших в предшествующий момент. Такое допущение хорошо "работает" применительно к движениям с небольшой амплитудой вертикального перемещения. А в цикле с большим вертикальным взмахом структура потока около крыла так слож..а, что в каждый данный момент она сильно зависит от предыдущей. Таким образом, допущение о независимости сил, действующих в последовательные моменты времени, нарушается, и в этом случае говорят о цикле, создающем нестационарное течение.
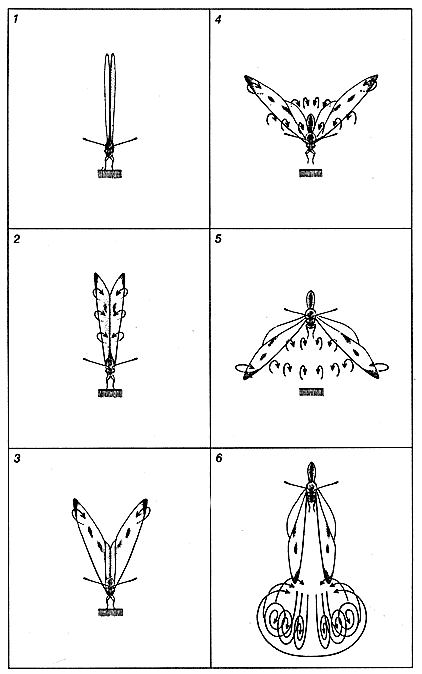
Один из механизмов, создающих вертикальную тягу в условиях нестационарного потока, описан Т. Вейс-Фогом из Кембриджского университета. В этом механизме, который носит название "флинг" *, основную роль играет ускорение циркуляции воздуха вокруг крыла (см. Weis-Fogh Т. Unusual Mechanisms for the Generation of Lift in Flying Animals, "Scientific American", November, 1975). В начале флинга крылья плотно сложены вертикально по отношению к спинке животного. Затем крылья расходятся, начиная с передних, ближайших к голове краев (см. рисунок справа). Расстояние между передними краями увеличивается по мере того, как крылья распахиваются вдоль линии, огибающей их задние края. Такое движение создает циркуляцию воздуха от нижних поверхностей крыльев вокруг их передних краев к расширяющемуся пространству между верхними поверхностями крыльев. Этот циркуляционный поток во время маха крыла вниз создает подъемную силу.
* Флинг (англ. fling) - резкое внезапное движение. - Прим. перев.Известно, что многие мелкие насекомые используют флинг, однако у более крупных видов он встречается редко. Как показал Эллингтон, если крылья большие, может произойти отрыв воздушного течения от верхней поверхности крыла, что приводит к резкому уменьшению подъемной силы. Однако если при этом крылья достаточно гибки, чтобы расхождение их передних краев происходило медленно, то течение не отрывается от крыла и достигается необходимая величина подъемной силы. Бабочки - самые крупные среди летающих организмов, способных к флингу, и по данным Эллингтона именно постепенное расхождение крыльев позволяет им пользоваться этим механизмом.
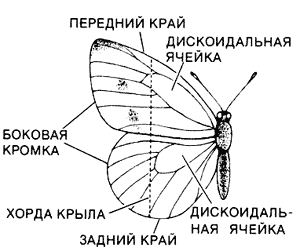
Флинг с распахиванием крыльев позволяет бабочкам рода Pieris подниматься в воздух. Этот процесс изображен справа на серии рисунков, частично основанных на данных Ч. Эллингтона из Кембриджского университета. Вначале крылья плотно сложены над телом (1). Флинг начинается с того, что передние края крыльев (около головы) отходят друг от друга (2). Процесс расхождения распространяется к заднему краю, и при этом возникает циркуляционный воздушный поток вокруг крыла (3). Когда расходятся задние края, крылья полностью отделяются друг от друга, начинается их мах вниз (4). Пока крылья движутся вниз, циркуляция воздуха около них продолжается, в результате чего создается подъемная сила (5). В конце маха, когда крылья оказываются в самом нижнем положении, циркуляционное течение отрывается от поверхности (6). После этого крылья поднимаются и начинается новый мах вниз. Строение крыла бабочки (слева) имеет большое значение для флинга. Жилкование таково, что передний край крыла жесткий, а вдоль хорды и по боковой кромке оно гибкое.Мои собственные предварительные исследования заставляют предполагать, что к гибкости крыльев, необходимой для медленного их распахивания, самое непосредственное отношение имеют крыловые жилки. Жилки образуют прочную опорную сеть, на которую натянута гибкая перепонка. У бабочек значительно меньше крыловых жилок, чем у других насекомых, особенно в дистальной (дальней от тела) части крыла. Наиболее крупные жилки отходят от основания крыла; среди них самая крепкая та, которая проходит по его переднему краю. Только в центральной части крыла, которая носит название дискоидальной ячейки, есть несколько жилок, перпендикулярных к жилкам, отходящим от основания крыла. Благодаря такому расположению жилок боковая кромка крыла весьма гибкая. Большая гибкость свойственна также крылу в направлении от переднего края к заднему (так называемая хордовая гибкость).По-видимому, жесткость переднего края и хордовая гибкость очень важны для распахивания крыльев. В самих крыльях мускулатуры нет и они двигаются за счет работы летательных мышц, находящихся в теле бабочки у основания каждую крыла. Со кращение этих мышц приводит в движение жесткий передний край крыла, и он совершает мах вниз. Благодаря хордовой гибкости остальной части крыла движение переднего края вызывает постепенное расхождение крыльев спереди назад. Так как боковая кромка крыла исключительно гибкая, имеет место также волна расхождения крыльев, распространяющаяся от вершины переднего края к телу насекомого.
Малое число жилок в крыле - необходимая предпосылка для процесса распахивания крыльев. В то же время такое жилкование исключает возможность использования жилок в качестве путей циркуляции жидкости для регуляции температуры тела. В устройстве крыла бабочки проявился общий принцип сочетания пользы и ущерба. Каждая из крупных анатомических структур бабочки принимает участие в осуществлении нескольких функций, и свойство, полезное для решения одной задачи, может быть лимитирующим фактором при решении другой. Создавая машину, от которой требуется выполнение нескольких функций, конструктор стремится путем компромисса достичь оптимального сочетания этих функций. По аналогии с машиной крыло бабочки можно рассматривать как пример наилучшего компромисса между несколькими важными функциями. Однако живые организмы создаются не по воле конструктора, а сами по себе, они эволюционируют, и вряд ли их следует считать оптимальными с инженерной точки зрения.
Есть и другое мнение, которое состоит в том, что анатомическое строение накладывает ограничения на способы осуществления жизненно важных функций организма, что эволюция и развитие происходят в неких рамках. Это можно продемонстрировать на примере бабочек, взяв для сравнения два гипотетических вида из рода Colias. Пусть эти виды отличаются от реальных тем, что обладают определенным адаптивным признаком в его крайних проявлениях Окраска крыльев у бабочек Colias обусловлена комбинацией двух пигментов - черного (меланина) и желто-оранжевого (птеридина). Конкретная, свойственная виду окраска зависит от количественного соотношения этих пигментов. Вместе с чешуйчатым покровом тела пигментация крыльев ограничивает выбор внешних условий, в которых бабочки-желтушки могут существовать.
Как я уже упоминал, бабочки рода Colias греются на солнце в латеральной позе, и крылья у них тем темнее, чем на большей высоте они живут. Поэтому представители гипотетического вида, обитающего на небольших высотах, должны иметь целиком желтые крылья и быть лишены чешуйчатого покрова, а у особей гипотетического вида, обитающего на больших высотах, должны быть полностью черные крылья и густой чешуйчатый покров. Если мы графически изобразим летные пространства этих двух гипотетических видов, то увидим, что они различаются гораздо сильнее, чем летные пространства реально существующих видов, которые я сравнивал между собой выше. Разница между ними максимальна з рамках ограничений, накладываемых двухпигментной системой окраски.
Конечно, условия внешней среды могут не укладываться в пределы, в которых возможна адаптация с помощью простой двухпигментной системы окраски крыльев. Чтобы представить себе виды, способные существовать в иных условиях, необходимо ослабить ограничения. Предположим, что к имеющимся двум пигментам добавлен еще третий, белый пигмент, а помимо существующих способов терморегуляции есть еще дополнительный - экспозиция с отражением солнечных лучей. Это расширяет нашу схему и позволяет войти в нее бабочкам poдa Pieris. Имея в распоряжении три пигмента и три позы, можно описать летные пространства всех видов бабочек семейства Pieridae, к которому относятся белянки и желтушки. По мере ослабления ограничений в рамки схемы попадают все более крупные таксономические группы. Если включить в рассмотрение некоторые биологические особенности, отсутствующие у Pieridae, например "разогревание" посредством интенсификации обмена веществ перед полетом и регуляцию тока гемолимфы, можно охватить всех представителей отряда Lepidoptera, к которому принадлежат дневные и ночные бабочки.
Таким путем можно вскрыть функциональный смысл таксономических категорий. Можно показать, что в ряде случаев таксоны различаются между собой по специфическим функциональным признакам, как, например, роды Pieris и Colias отличаются друг от друга окраской крыльев и позами. Поскольку признаки, подобные окраске крыльев, генетически обусловлены, расшифровка системы ограничений проливает свет и на эволюционные отношения между организмами. Скажем, инженерный анализ позволяет нам представить, как бы повлияла мутация, в результате которой возник третий пигмент у бабочек рода Colias, на диапазон местообитаний этих бабочек. Сочетая при изучении живых организмов инженерный подход с анализом эволюционных отношений, можно подойти к пониманию того, как физические механизмы ставят пределы эволюционным изменениям, как, в частности, сложилась та хрупкая конструкция, которая позволяет бабочке порхать в воздухе.
Литература
J.G. Kingsolver and T.L.Daniel. Mechanical determinants of nectar feeding strategy in hummingbirds: energetics, tongue morphology, and
licking behavior. // Oeco/ogia, Vol. 60, pages 214-226; 1983.J.G. Kingsolver, Thermoregulation and flight in colias butterflies: elevational patterns and mechanistic limitations. // Ecology, Vol. 64, pages 534-545; 1983.
C. P. Ellington, The aerodynamics of hovering insect flight. // Philosophical Transactions of the Royal Society, B. Biological Sciences, Vol. 305, No. 1122, pages 1-181; February 22, 1984.

Май 2005