№12, 2003 г.
© В.В. Алёшин, Н.Б. Петров
Условно нейтральные признаки В.В. Алёшин, Н.Б. Петров
Владимир Вениаминович Алёшин, к.б.н., вед. научн. сотр. отдела эволюционной биохимии
Научно-исследовательского института физико-химической биологии им.А.Н.Белозерского
Московского государственного университета им.М.В.Ломоносова.Николай Борисович Петров, д.б.н., зав. лаб. геносистематики животных того же института.
В нынешней теории эволюции есть одно кажущееся противоречие: непрерывная эволюция приводит к дискретному результату - новым видам. Вряд ли кто-то станет спорить, что это следствие вымирания. По Ч.Дарвину, борьба за существование заставляет возникающие формы уходить от конкуренции, приспосабливаться к различным условиям, и это ведет к дивергенции. Так что виды должны отличаться в первую очередь по адаптивным признакам. Однако тот, кто хоть однажды знакомился с разнообразием видов не по Красной книге, где собраны представители с броской внешностью, знает, как трудно бывает определить видовую принадлежность найденного экземпляра. Озадачивает, что в определительных ключах почему-то указаны признаки, функциональное назначение которых часто непонятно. Неужели только такой узор на крыльях бабочек или раковин морских моллюсков конусов, форма зазубринок по краю листовой пластинки и длина шипов на пыльцевых зернах - основной результат дивергенции и только они обеспечивают выживание вида в природе? А если бы они были чуть-чуть иными?
Нередко диагностические признаки имеют двойственную природу: и приспособительную, и как бы случайную. Так, шипики в жевательном желудке короедов нужны для превращения древесины в опилки. Отличия в вооружении желудков у разных видов тоже понятны. Ведь исходная прочность древесины, к которой специализированы короеды, не одинакова: одному требуется “рашпиль”, а другому - мелкозубчатый “напильник”. Но почему рельеф желудка неповторим у десятков видов? А нередко видоспецифические орнаменты покрывают и поверхности, не используемые в качестве абразивных. Ихтиолог Л.С.Берг, адепт законов, направляющих эволюцию, в качестве структур непонятного назначения приводил выросты переднеспинки у небольших насекомых горбаток (рис.1) [1]. На самом деле выросты делают горбаток колючими. Так что их полезность сомнения не вызывает. Но почему они именно такой формы и настолько отличаются у разных видов?
Рис. 1. Формы, возникновение которых, по мнению Л.С. Берга.,
“трудно объяснить <…> естественным отбором” [1].Неясность функционального значения многих диагностических признаков породила диаметрально противоположные мнения, разбила специалистов на две партии. Одну образовали, условно говоря, ортодоксальные дарвинисты, считающие, что мы просто не понимаем смысла видовых различий, на самом деле важных и безусловно приспособительных. На другом полюсе собрались ревизионисты, утверждающие, что эволюция идет не по Дарвину. Это позиция была очень соблазнительна: открытие эволюции органического мира стало важным и эмоционально насыщенным событием в биологии XIX в., повлиявшим на культурную жизнь людей. Заново открыть эволюцию, конечно, невозможно, но многим хотелось найти или придумать хотя бы свой ее главный фактор.
Итак, что заставило виды отлиться именно в те формы, которые мы наблюдаем: неизбежная закономерность эволюции или случайность? Всякое ли эволюционное изменение приспособительно? На эти старинные вопросы трудно дать достаточно обоснованные ответы, если ограничиваться традиционными аргументами спорящих более века сторон.
Нейтральная эволюция и молекулярные часы
Иногда биологам приходится иметь дело не с целыми организмами, а с их фрагментами (при изучении питания - по содержимому желудков, фауны и флоры недавнего прошлого - по захоронениям в торфе и т.п.). Такие фрагменты или целые организмы (личинки, самки) под микроскопом чаще всего можно определить только до рода, потому что видовые признаки не обнаруживаются, а внешний вид и анатомическое строение большинства органов у близких видов совершенно неотличимы даже для специалиста. Совсем иная картина открылась при изучении макромолекул. Хотя обмен веществ у всех живых существ сходен, белки-ферменты, катализирующие в клетке одну и ту же химическую реакцию, у разных видов почти всегда хоть немного, но отличаются по первичной структуре. В сущности наибольшее биоразнообразие открылось с изучением макромолекул. И оказалось, вызвано оно не движущим естественным отбором, а игрой случая.
Среди 20 типов аминокислот, входящих в состав белков, некоторые химически сходны. Легко представить, что замена аминокислоты на похожую мало повлияет на общие свойства белка, состоящего из сотен аминокислотных остатков. Конечно, это не относится к остаткам, находящимся в активном центре фермента: они одинаковы не только у близких видов, но и у всех живых существ. Но даже самые консервативные гены изменчивы в силу вырожденности генетического кода.
Проверить, влияют или не влияют на приспособленность морфологические различия близких видов, почти невозможно, а вопрос о нейтральности молекулярных решается экспериментально. Так, чужеродный ген, введенный в бактерии или дрожжи методами генной инженерии, нередко полностью компенсирует дефект собственного гена, по крайней мере в лабораторных условиях. В экспериментах с многоклеточными также показана возможность полноценной неродственной замены, в том числе и генов, управляющих эмбриональным развитием [2].
Если не движущий отбор, то какая сила вызвала эволюцию вариабельных участков, состав которых не важен для работы фермента? Предположим, некоторые аллельные состояния гена мало влияют на выживание особи в довольно разнообразных условиях. Это допущение не противоречит бытовым наблюдениям над людьми с разным фенотипом, генно-инженерной практике или ящичным экспериментам с мутантными линиями плодовых мушек. Вопрос в том, может ли нейтральный признак распространиться в популяции, т.е. победить исходный аллель.
Выживание и размножение особи в природе зависит не только от абстрактной величины - адаптивности, но и от случая. Отбор предпочитает более приспособленных, однако случай может дать шанс кому угодно, просто шанс этот меньше. Если приспособленность двух аллелей почти не отличается, то их соотношение в популяции будет колебаться по случайным причинам. Впервые такие колебания частот нейтральных аллелей предсказали в 1931 г. Н.П.Дубинин, Д.Д.Ромашов и С.Райт. Ситуация аналогична игре в орлянку (или на бирже): хотя вероятность проигрыша одинакова при конкретном броске (1/2), тем не менее, если два человека с ограниченной суммой долго играют между собой, один непременно проиграет. Рано или поздно (тем скорее, чем меньше денег) ему не повезет, а отыграться он не сможет, потому что нечего будет поставить на кон. В популяционной генетике этому соответствует полная утрата одного аллельного варианта [3]. Одинаковая приспособленность проявляется в том, что невозможно предсказать, какой аллель (или какой игрок на бирже) победит. Время от времени это будет какой-либо вновь возникший мутант. Игра случая при достаточно продолжительной независимой эволюции приведет к накоплению отличий в функционально несущественных областях генов. Поскольку они нейтральны, любые из них одинаково вероятны и у каждого вида будут независимыми, а различия составят сумму мутаций, зафиксированных в эволюции после их дивергенции от общего предка.
Не так легко представить фактор, который бы сдерживал или, наоборот, стимулировал распространение в популяции нейтральных мутаций. Согласно простейшей гипотезе Э.Цукеркандля и Л.Полинга, нейтральные мутации возникают и закрепляются с относительно постоянной скоростью (вероятностью). Чем дольше независимая эволюция видов, тем больше мутаций успеет закрепиться. Их накопление оказывается своего рода молекулярными часами, по которым можно оценить время независимой эволюции видов и построить филогенетическое дерево *.
* Здесь не важно, насколько точны молекулярные часы [4]. По крайней мере по сравнению с радиоуглеродными часами они менее точные хотя бы потому, что статистические закономерности, лежащие в их основе, проявляются не на миллиардах атомов, а всего на тысячах нуклеотидных остатков, если учитывать какой-нибудь один ген.В неорганическом мире разрушение любого сложного предмета или системы можно предотвратить, только затрачивая энергию на их поддержание (консервацию или ремонт). Без ремонта тепловое движение молекул, геологическое выветривание, космические лучи и другие энтропийные факторы порождают и постепенно увеличивают хаос в сложной системе. Биологические виды удерживают сложную морфологию, принося в жертву энергию, заключенную в телах погибших и не оставивших потомства мутантов. Где алтарь стабилизирующего отбора угасает, там энтропийные факторы (мутационный процесс) в скором времени разрушат сложную форму. Так происходит редукция органов, ставших бесполезными (глаза пещерных рыб и т.п.). Наличие в макромолекулах неизменных для всех царств функциональных участков наряду с вариациями, отличающими близкие виды, определяется совместным действием энтропийных (мутагенез) и антиэнтропийных (отбор) факторов.В целом эволюция макромолекул с ее очевидным нейтральным компонентом, молекулярными часами и малоприметностью движущего отбора кажется более понятной, но совсем непохожей на эволюцию макроскопических признаков, по которой выведены закономерности эволюции, записанные в учебниках.
Прерываемое равновесие
Вторая, после теории нейтральной эволюции, идея, взбудоражившая научное сообщество и вызвавшая шквал откликов, хвалебных и ругательных, пришла из палеонтологии. Н.Элдридж и С.Гулд, изучая окаменевшие остатки моллюсков, захороненных на месте пресноводного озера в Восточной Африке, описали характерную для палеонтологии ситуацию: в достаточно больших по толщине слоях осадков они обнаруживали раковины неизмененной формы. Вдруг, начиная с какого-то горизонта, прежняя форма навсегда исчезала, а вместо нее возникала похожая, но все-таки отличающаяся (как бы новый вид моллюска из того же рода). И это наблюдалось много раз по всей толще разреза. Поскольку переходных форм палеонтологи не находили, они предположили, что виды сами по себе очень стабильны, но иногда стабильность почему-то прерывается, и скачком, без постепенных изменений, возникает новый вид. Такой способ эволюции Элдридж и Гулд назвали прерываемым равновесием. Сторонники этой идеи образовали лагерь пунктуалистов (от англ. punctuate - прерывать), объявили ее новым словом науки, а своих противников, градуалистов, - ретроградами. Градуалисты в свою очередь не оставались в долгу. Действительно, вряд ли скорость осадконакопления в разрезах была неизменна и отсутствовала эрозия, уничтожавшая время от времени осадки, содержавшие переходные формы. Невероятно, чтобы на протяжении всех геологических эпох ископаемый водоем был замкнут и в него не могли вселяться иммигранты из соседних водоемов, где они возникли и где оставили ископаемые переходные формы. Но главный аргумент градуалистов заключался в ясности идеи отбора как естественного, постепенно действующего фактора эволюции против туманных причин стазиса и его прерывания в трактовке пунктуалистов.
Дискуссия вокруг идеи прерываемого равновесия показала необходимость переоценки давно известных фактов неравномерности морфологической эволюции. А то, что мы знали об эволюции ДНК, в основном ограничивалось градуальной теорией нейтральной эволюции и эмпирическим обобщением молекулярных часов, идущих с постоянной скоростью. В этих теориях не было места ни стабильности, ни ее неожиданному прерыванию.
Прерываемое равновесие в эволюции макромолекул
У всех организмов, от бактерии до человека, рибосомная РНК выполняет одинаковую функцию - участвует в синтезе белка. Одни ее участки почти неизменны, другие изменчивы по составу, но образуют консервативные элементы вторичной структуры, например короткие внутримолекулярные двойные спирали, соединенные уотсон-криковскими парами. Если нуклеотид в одной ветви меняется, то для сохранения спирали меняется и противолежащий - комплементарный ему [5]. Конечно, компенсация возникает не из-за целесообразности мутаций. Просто нейтральными (а только они могут сохраниться) оказываются двойные мутации.
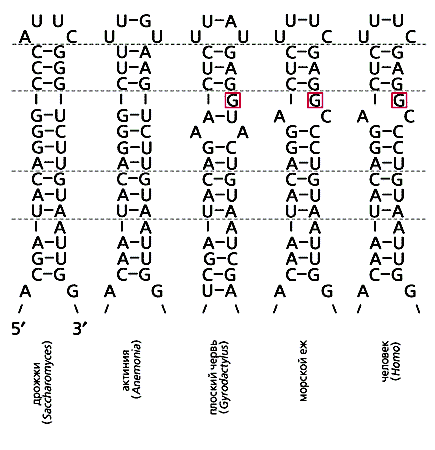
Один из элементов рРНК, изменчивый по составу, но консервативный по вторичной структуре, - спираль с порядковым номером 17, она представлена в живой природе двумя основными формами. Одна, длиною 13 нуклеотидных пар, почти универсальна (обычна у водорослей, высших растений, жгутиконосцев, грибов, низших животных: губок и кишечнополостных). Вторая выявлена у двусторонне-симметричных животных (Bilateria) и отличается вставкой неспаренного, “лишнего”, нуклеотида, который располагается в одном и том же месте в моделях этой спирали (рис.2). Спираль из 131/2 пар почти столь же эволюционно стабильна, как из 13 пар, и распространена у позвоночных, членистоногих, моллюсков, иглокожих, плоских, кольчатых червей и других животных. Прежнее равновесие оказалось прервано и установилось новое.
Рис.2. Модификации спирали 17 рРНК у двусторонне-симметричных животных. Структура этой спирали у кишечнополостных сохраняет предковое состояние, подобное таковому у грибов (две модели слева). Пунктирные линии отделяют гомологичные участки для удобства сопоставления. Цветом выделен индикаторный признак (специфический нуклеотид) спирали, показывающий монофилию двусторонне-симметричных животных.Очевидно, дополнительный нуклеотид внедрился в ген рРНК общего предка Bilateria и унаследовался современными типами. Поскольку его нет в рРНК губок, гребневиков, кишечнополостных, то общий предок Bilateria возник уже после отделения от “радиальных” животных. Филогенетика, конечно, не указывает прямо, каким образом он произошел и как был устроен (была ли у него паренхима, как у плоских червей и личинок кишечнополостных, или все его ткани были эпителизованы, как у высших многоклеточных и взрослых кишечнополостных). Это выходит за область ее применения. Но сходная генетическая основа для развития осевых структур в онтогенезе насекомых, позвоночных и плоских червей говорит о том, что ближайший общий предок Bilateria был двусторонне-симметричным, а не радиальным [6].Наличие альтернативных состояний в структуре спирали 17, каждое из которых стабильно, напоминает состояние стазиса и его нарушения в моделях пунктуалистов. Примерно так же меняется конфигурация и других спиралей в рРНК: например, укорочение спирали 42 (оно помогло расшифровать родственные отношения низших многоклеточных), локальные нарушения комплементарности в спирали 49 и др. [7].
Закон Долло для макромолекул
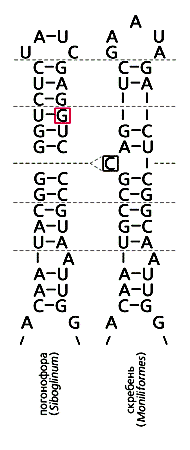
Характерная особенность спирали 17 Bilateria, неспаренный нуклеотид, отсутствует в рРНК погонофор и вестиментифер - глубоководных свободноживущих червей, у которых нет пищеварительной системы. Вестиментиферы прославились симбиозом с хемотрофными бактериями, извлекающими химическую энергию из горячих высачиваний “черных курильщиков”, обеспечивая в глубинах океана синтез органических веществ для себя и всего сообщества [8]. Погонофоры таким же образом используют холодные высачивания природного газа метана. У этих червей для “лишнего” нуклеотида нашелся парный, и их спираль состоит из 14 пар. Погонофоры и вестиментиферы - близкие родственники, и своеобразное устройство спирали 17 в рРНК еще одно подтверждение их монофилии (рис.2).
Другие животные без кишечника, паразитические колючеголовые черви или скребни (Acanthocephala), потеряли типичный “лишний” нуклеотид, но приобрели вместо него свой, специфический. Он у них пиримидин, а не пурин и находится в 5’-, а не в 3’-ветви спирали 17. Круглые черви на первый взгляд “вернулись” к предковому состоянию спирали: у них нет неспаренного нуклеотида и общая длина спирали 13 пар. Но если у растений, грибов, протистов и “радиальных” животных в четвертой паре от вершины в 3’-ветви почти всегда находится пиримидин (обычно остаток тимидина), то у нематод - пурин (гуанин, реже аденин), как положено “лишнему” нуклеотиду Bilateria, номер которого - четвертый. Похоже, круглые черви избавились не от “лишнего”, а от следующего за ним нуклеотида.
Спираль 17 своеобразно устроена и у бескишечных ресничных червей отряда Acoela, пресноводных брюхоресничных червей, мизостомид, тихоходок и некоторых других животных [9]. Среди миксоспоридий имеется даже несколько ее вариантов. Складывается впечатление, что эта структура следует известной шутке: менять конфигурацию нельзя, но если очень хочется, то можно. В результате она принимает какую угодно форму, но постоянную в рамках монофилетической группы (типа, отряда, группы видов). Вероятно, с увеличением числа изученных видов найдутся сходные модификации у неродственных видов. Например, как и у погонофор и вестиментифер, устроена она у бокоплавов - единственного отряда ракообразных, где произошла такая же конвергентная достройка пары к “лишнему” нуклеотиду.
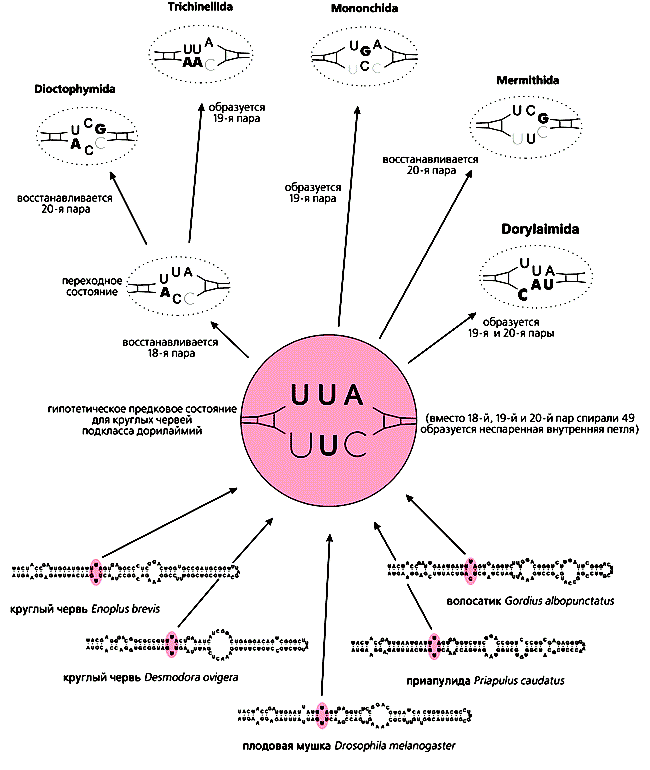
Подобная специфичность обнаруживается и в спирали 49 рРНК. Так, у многих видов в определенном участке вместо комплементарной уотсон-криковской пары закономерно находятся два неспаренных пиримидина. Но у пяти отрядов круглых червей Dorylaimia они или смещены с 19-й пары на 18-ю или 20-ю, или окружены необычными нуклеотидами, притом современные последовательности нельзя легко вывести одну из другой (рис.3). Значит, уже у их общего предка нарушилась комплементарность соседних пар, а затем произошла полная или частичная компенсация (в разных отрядах независимо, за счет либо обратных, либо компенсаторных мутаций). Стабилизировать одну или две пары из трех можно 18 способами, и не удивительно, что в пяти отрядах Dorylaimia все они разные.
Рис.3. Эволюция внутренней петли спирали 49 малой рРНК у круглых червей подкласса дорилаймий. Цветом показано, что на месте 19-й пары находятся два противолежащих нуклеотида - пиримидина, ограниченных уотсон-криковскими, 18-й и 19-й парами. (Rusin L.Y. et al. // Nematology. 2003.)Сложность макромолекул (РНК и белков) не меньше, чем морфологических структур и органов. Множество конструктивных способов решения задач, диктуемых отбором (например, стабилизировать одну из спиралей рРНК), объясняет многообразие их реализации (при условии случайности мутаций). Так, если какое-то предыдущее изменение потеряло адаптивность, то возврат в исходное состояние только одна из многих возможностей, и весьма вероятно, что вместо возврата появится что-то новое. Этот принцип необратимости эволюции, названный по имени бельгийского ученого, сформулировавшего его, законом Долло, на уровне макромолекул получает ясное и наглядное объяснение как результат стабилизирующего отбора на фоне случайных мутаций.Приспособление или компенсация?
В основу современной парадигмы молекулярной эволюции легли две тесно связанные гипотезы: концепции значительного нейтрального компонента в эволюции макромолекул и молекулярных часов. Постоянство мутационного процесса и случайный характер размножения обладателей различных аллельных вариантов обеспечивают весьма быстрые, безотносительные к движущему отбору, изменения в митохондриальных геномах, межгенных участках, интронах, вариабельных областях рибосомных генов, отличающие даже близкие виды. Напротив, в спиралях 17 и 49 малой рРНК обычно фиксируются только те мутации, которые не затрагивают вторичную структуру, пока не возникнет временная их нестабильность, результат которой - измененное состояние - далее вновь стабильно наследуется. По сути спирали 17 и 49 эволюционируют в режиме прерываемого равновесия.
При наличии в белках и рРНК активного центра и “неважных” участков, переменная эволюционная консервативность несколько неожиданна. Тем не менее такие участки небесполезны: они должны сблизить функционально активные остатки на строго определенное расстояние, ориентировать их под нужными углами друг к другу, облегчить доступ в активный центр субстратов и удалить продукты, заякорить макромолекулу в нужном отделе клетки, связать сигнальные молекулы и, конечно, при этом не затрагивать ничего постороннего. Многие эволюционно устойчивые состояния рРНК отличаются вставками или делециями отдельных нуклеотидов, т.е. мутациями, локально изменяющими вторичную структуру. Можно предположить (по аналогии с комплементарными парами в спиралях), что они, дабы не повлиять существенно на структуру рибосомы и не снизить приспособленность, должны сопровождаться одновременной компенсацией во взаимодействующей молекуле (РНК или полипептиде).
100 лет назад английский философ и эволюционист Г.Спенсер понял, что одновременные согласованные мутации очень редки, если они независимы. Тогда еще не знали об эмбриональной индукции, благодаря которой в онтогенезе целые комплексы структур меняются согласованно в зависимости от единственной мутации, влияющей на развитие индуктора. Спенсер полагал, что для наблюдаемой скорости эволюции необходима определенная (а не дарвиновская неопределенная) изменчивость. Молекулы, в отличие от органов, изменяются независимо, так что длительные периоды стазиса в их эволюции (до случайного изобретения новой гармонии) неудивительны.
Отсутствие способов компенсации делало бы любые функциональные участки генов исключительно консервативными, а мутационный груз очень тяжелым. Один из таких способов показывает спираль 49: она достаточно длинная, и одна-две мутации, нарушающие комплементарную пару, приведут только к локальному изменению и скорее всего будут нейтральными или слабо вредными. Но любое следующее нарушение, затрагивающее еще одну пару, вызовет “фазовый переход” - расплетение спирали в целом и гибель мутанта. Теперь фиксироваться смогут только компенсаторные мутации. Значит, используя предшествующий запас прочности, можно изменить одну комплементарную пару в спирали за две мутации, предварительно исчерпав запас прочности: доведя спираль почти до полного развала и пожертвовав всеми последующими мутантами без компенсации. Несовершенная короткая спираль вряд ли долговечна. Она должна быстро стабилизироваться в результате обратной или компенсаторной мутации и предстать либо в виде неизмененного исходного состояния, либо в виде двойной мутации.
Но спираль 49 вовлечена и в межмолекулярные взаимодействия. Ее 3’-ветвь комплементарна не только 5’-ветви, но и так называемой транспортно-матричной РНК [10]. Благодаря этому спираль может расплестись и связать тмРНК, что необходимо в некоторых “аварийных” ситуациях *. Следовательно, чтобы мутации в этой области были нейтральными, они должны компенсироваться не только в комплементарной ветви спирали, но и в тмРНК. Пока тмРНК найдена только в клетках бактерий, но неравномерный характер эволюции спирали 49 эвкариот заставляет думать, что и у них нуклеотиды этой спирали участвуют в межмолекулярных взаимодействиях.
* Впервые с действием тмРНК столкнулись генные инженеры, когда с удивлением обнаружили, что различные неправильно сконструированные гены (без стоп-кодона) приводят к синтезу в бактериальной клетке белков с одинаковыми C-концевыми аминокислотами, не закодированных к тому же в искусственных генах. Если мРНК по какой-то причине лишена стоп-кодона (а мРНК без стоп-кодона постоянно возникает в клетке, поскольку эта молекула не вечна и постепенно разрушается), то бактериальная рибосома не прерывает сразу трансляцию, а дожидается, когда с ней свяжется тмРНК, на которую трансляция и переключится. В тмРНК есть стоп-кодон, а перед ним несколько триплетов, кодирующих аминокислотные остатки - мишени внутриклеточных протеаз. Поэтому дефектный белок, синтезированный с неполной матрицы, будет в клетке очень быстро распознан и расщеплен.Таким образом, мутации, требующие компенсации, возникают редко (относительно времени существования филогенетических линий), не подвержены реверсиям (по закону Долло), и поэтому те из них, которые возникли в стволовой группе, маркируют ветви филогенетического дерева [7]. Их нельзя рассматривать как адаптивные: нет оснований думать, что это приспособления к каким-то новым особенным функциям. В то же время скорость их фиксации не такая, как у нейтральных мутаций: их возникновение невозможно предсказать, исходя из времени существования филогенетической линии по результатам калибровки молекулярных часов.Такие мутации, названные нами условно нейтральными, очень важны для филогенетики [11]. Действительно, как распознать древнюю линию, отдельные ветви которой существуют, например, с кембрия? Их общие признаки не должны разрушиться нейтральными мутациями. Значит, они адаптивны? Но в чем состоит адаптивное значение различий в макромолекулах, выполняющих неизменные клеточные функции? И если они адаптивны, возникает новый вопрос о вероятности конвергенции. Может, признаки типов и классов эволюционируют по другим законам, нежели нейтральные признаки низших таксономических уровней, и молекулярная биология наконец-то нашла пресловутую границу между микро- и макроэволюцией? Представление об условной нейтральности этих признаков снимает остроту всех перечисленных вопросов, а заодно служит наглядной моделью прерываемого равновесия, более понятной, чем в случае морфологической эволюции.
Совместное действие на генетический аппарат стохастического мутационного процесса и стабилизирующего отбора вносит в эволюцию функционально значимых признаков компонент неравномерности, своеобразной квантованности и непредсказуемости их изменений во времени, тогда как нейтральные признаки меняются постепенно, пропорционально времени независимой эволюции, но направление их изменения принципиально непредсказуемо (в этом и проявляется их нейтральность). Таким образом, биологическая эволюция включает как содержательную, так и временну?ю неопределенность, и вряд ли надо требовать от теории эволюции такой прогностичности, как от ньютоновской механики.
Наследственные болезни и условно нейтральные мутации
Наследственная болезнь - это в простейшем случае такая мутация, которая приводит к замене одной из аминокислот в кодируемом белке, изменяет или дестабилизирует его пространственную структуру. В результате белок перестает выполнять свою функцию или выполняет ее хуже. Например, патологически измененный гемоглобин хуже или, наоборот, чересчур прочно связывает кислород или плохо сохраняется в эритроцитах - одним словом, это нарушает его транспортную функцию и приводит к анемии. Вредные мутации не могут фиксироваться в популяциях по законам нейтральной эволюции, и можно было ожидать, что те аминокислотные замены, которые патогенны для человека, не будут обнаружены у других видов. Однако А.С.Кондрашов с соавторами [12] обнаружил, что такие мутации встречаются как видовые признаки, хотя и намного реже, чем мутации, не попавшие в список патогенных для человека, причем примерно с одинаковой частотой у приматов, рыб или земноводных, т.е. независимо от родства с человеком. Может быть, они потому фиксируются редко, что портят белки других организмов, как и белки человека? Но тогда как они вообще фиксируются? Почему макаки и ксенопусы с патогенной для человека мутацией не болеют?
Вредная мутация может стать нейтральной, если одновременно в структуре макромолекулы (этой же или другой, но взаимодействующей с ней в клетке) происходит изменение, компенсирующее дефект. Одна такая пара замен обнаружена в глобине [12], причем мутантные аминокислоты разнесены в последовательности, но в нативной молекуле пространственно сближены. Вероятность двух одновременных мутаций крайне низка, поэтому частота появления жизнеспособных двойных мутантов очень мала и зависит только от одновременного возникновения двух таких мутаций. Нейтральные изменения между соответствующими генами человека и ксенопуса или рыбы мало повышают возможность фиксации условно вредной мутации, поскольку “компенсирующая” мутация сама по себе вряд ли нейтральна и скорее всего тоже может существовать только в паре с уже известной вредной мутацией.
Унификация повторяющихся последовательностей
Для полноты картины рассмотрим еще один пример, когда диалектика случайных мутаций и случайного размножения приводит к единообразию внутри вида при межвидовых отличиях по нейтральным признакам. Возьмем гены рибосомной РНК. В эвкариотическом геноме их обычно 200-300, сгруппированных в несколько больших тандемных кластеров по способу “голова к хвосту”. Много копий необходимо, чтобы обеспечить нужный уровень транскрипции, ведь рРНК - одна из самых массовых макромолекул в клетке. По канонам классической генетики, ее гены, расположенные в различных локусах, должны накапливать мутации и эволюционировать независимо. В результате в одном геноме должны бы присутствовать потомки первой дупликации генов рРНК (произошедшей у самого первого эвкариота), которые должны отличаться между собой не меньше, чем отдельные гены рРНК амебы и человека! Однако члены семейства генов рРНК похожи друг на друга, как близнецы. Это очень кстати, иначе возникли бы исключительные трудности при сборке рибосом, в состав которых, кроме четырех различных рРНК, входит несколько десятков белков, и все должны быть подогнаны друг к другу!
Оказалось, нуклеотидные последовательности в тандемных блоках эволюционируют согласованно [13, 14]. Конечно, ничто не может помешать возникновению мутации в любом локусе. Вопрос в ее дальнейшей судьбе. Либо мутантный аллель, либо его сосед вскоре дуплицируется (в блоках это происходит часто из-за неравного кроссинговера при мейозе, а также по механизму генной конверсии и транспозиции с перемещением на другие хромосомы, только с меньшей частотой). Но неограниченное размножение одного гена в геноме невозможно: кроме дупликаций бывают делеции, и если они не снижают число рибосомных генов ниже нормы, то их обладатели сохраняются в популяции. В результате в геноме число рибосомных генов то немного возрастает, то убывает, и при этом меняется соотношение исходного и мутантных вариантов гена рРНК. Когда-то частота одного из них станет равна единице. Если новый вариант рибосомы не хуже исходного, то на втором этапе, как учит теория нейтральной эволюции, он может зафиксироваться не только в геноме, но и в популяции [3].
Таким образом, мутационному процессу противостоят унифицирующие факторы: неравный кроссинговер при образовании гамет (сам по себе мутационный фактор), гибридизация и случайное размножение особей в популяции. Конечно, унификация - не закон природы, а только результат соотношения различных факторов (частоты мутаций и неравного кроссинговера, селективности мутантных вариантов, числа элементов в тандемном блоке, тандемов в хромосоме, хромосом с генами рРНК, частоты переноса аллелей с одной хромосомы на другую, эффективной численности популяции). Если бы значения всех этих параметров были известны, их можно было подставить в формулу и предсказать, какой процесс будет в данном случае преобладать: расхождение копий в геноме или их унификация. За очень редкими исключениями индивидуальные аллели рРНК почти не отличаются в пределах одного генома (и одного скрещивающегося сообщества, т.е. биологического вида). Вместе с генами рРНК унифицируются и соседние фрагменты ДНК, которые вырезаются и не попадают в рибосому. Их функция ничтожна (не мешать созреванию рРНК), тем не менее они так же стабильны внутри вида, как и гены рРНК [13]. Это результат унификации. Но, естественно, самые разнообразные мутации таких участков не вредят их функции. Благодаря мутационному процессу и нейтральной эволюции они довольно сильно отличаются даже у близких видов.
Таким образом, половой процесс и гибридизация - главный фактор унификации, а вовсе не источник разнообразия, за который его принимают по недоразумению, поверив селекционерам на слово. Описанный “молекулярный привод” обеспечивает единство скрещивающегося сообщества по множеству тандемно сгруппированных повторяющихся последовательностей и его скачкообразное отличие от других биологических видов. Оно выявляется с помощью различных методик [15] и позволяет установить биологические виды только по коллекционным экземплярам, без наблюдений за скрещиванием в природе. Этот относительно понятный, постепенно действующий механизм градуальной эволюции создает прерывистость нейтральных генетических маркеров, в точности напоминающую прерывистость морфологических диагностических признаков.
Литература
1. Берг Л.С. Номогенез, или эволюция на основе закономерностей / Тр. по теории эволюции. 1922-1930. Л., 1977. С.95-311.
2. Ronshaugen M. et al. // Nature. 2002. V.415. ?6874. P.914-917.
3. Кимура М. Молекулярная эволюция: теория нейтральности. М., 1985.
4. Антонов А.С. Эволюция генов растений: вызов теории “молекулярных часов” // Природа. 1986. №7. С.68-77.
5. Woese C.R. et al. // Microbiol. Rev. 1983. V.47. №4. P.621-669.
6. Воронов Д.А. Старая гипотеза “перевернутости” хордовых подтверждается // Природа. 2000. №11. С.18-22.
7. Алёшин В.В., Петров Н.Б. Регресс в эволюции многоклеточных животных // Природа. 2001. №7. С.62-70.
8. Малахов В.В. Вестиментиферы - автотрофные животные // Сорос. образоват. журн. 1997. №9. С.18-25.
9. Алёшин В.В. и др. // Молекуляр. биология. 1999. T.33. №2. С.319-329.
10. Muto A. et al. // Trends Biochem. Sci. 1998. V.23. №1. P.25-29.
11. Петров Н.Б., Алёшин В.В. // Генетика. 2002. T.38. №8. С.1043-1062.
12. Kondrashov A.S. et al. // Proc. Natl. Acad. Sci. USA. 2002. V.99. №23. P.14878-14883.
13. Доувер Г. и др. Динамика эволюции геномов и дифференциация видов // Эволюция генома. М., 1986. C.329-356.
14. Ратнер В.А. и др. Проблемы теории молекулярной эволюции. Новосибирск, 1985.
15. Медников Б.М., Шубина Е.А., Мельникова М.Н. Молекулярные механизмы генетической изоляции // Природа. 2001. №5. С.40-47.