№10, 2004 г.
© Акифьев А.П.
Избыточная ДНК - генетическая квадратура круга?
А.П. Акифьев
Алексей Павлович Акифьев, д.б.н., проф., гл. научн. сотр., рук. группы мутагенеза Института общей генетики им.Н.И.Вавилова РАН
Главный парадокс генома высших эукариот
Многие биологи полагают, что понять истинный смысл структур и функций систем живых клеток и организмов можно только изучив причины и способ возникновения их в эволюции. В середине ХХ в., так же как и во времена Ч.Дарвина, в учебниках биологии схематически изображалось развитие жизни на Земле в виде эволюционного древа. В его подножии находились простые и, как думали, самые древние организмы, а на вершине непременно располагался человек. Поэтому была вполне естественной точка зрения, согласно которой у Homo sapiens должно быть наибольшее число генов. А поскольку генетический материал - это ДНК, то следовало ожидать, что и по ее количеству человек непременно должен быть рекордсменом среди форм той единственной белково-нуклеиновой жизни, которая существует на Земле (а может быть, и вообще в Космосе).
Однако еще в конце 40-х-начале 50-х годов (т.е. до создания модели двойной спирали) ученые, в частности американские биохимик А.Мирский и цитолог Х.Рис, абсолютно четко доказали, что стоящий на вершине эволюции вид H.sapiens обладает не самым крупным геномом [1]. (Геном - это количество ДНК, содержащееся в гаплоидном, одинарном, наборе хромосом.) Если соединить конец в конец 23 молекулы ДНК, составляющих гаплоидный набор человека, получится нить длиной около 1 м. (Длина ДНК самой крупной первой хромосомы человека составляет около 7.5 см.) Но геномы саламандры и лилии почти в 30 раз превышают размер человеческого генома. Налицо полный крах представлений о соответствии сложности организма и величины его генома. Это явление, даже получившее собственное название - парадокс размера генома (C-value paradox), до сих пор не объяснено. Нет ответа и на такие вопросы, как: почему весьма мало различаются размеры генома человека и мыши; почему геномы хвостатых амфибий варьируют в пределах двух порядков, а бесхвостых - довольно близки по размерам.
В настоящее время расшифрованы полные геномы (т.е. нуклеотидные последовательности ядерной ДНК) нескольких организмов, в том числе, разумеется, и человека. Оказалось, что в сумме его гены вместе с регуляторными участками едва ли превышают 3-5% всего генома. О назначении остальной ДНК сегодня, как и 50 лет назад, мы фактически ничего не знаем и не понимаем ни ее эволюционной роли, ни механизма происхождения. Конечно, можно формально допустить, что в ходе эволюции размер генома сменяющих друг друга видов не оставался постоянным. Но каким образом геномы могли быстро увеличиваться и, что более загадочно, столь же стремительно уменьшаться?
По отношению к геномному парадоксу в англоязычной научной литературе часто используются термины “enigma” и “puzzle”. Оба они означают “загадочный”, но второй термин можно перевести и как “головоломка” и даже “неразрешимый вопрос”. То и другое не противоречит смыслу парадокса. К нему вполне применимо также древнее понятие “квадратура круга” (но только генетическая), поскольку задача как таковая существует, но пути ее решения традиционными способами не просматриваются.
Биологическая роль избыточной ДНК остается одной из главных тайн эволюции и, вероятно, наиболее трудной проблемой эволюционной генетики высших организмов. Тем не менее уже есть некоторые экспериментальные результаты и наблюдения конкретных клеточных процессов, на основе которых можно попытаться объяснить геномный парадокс. Об этом и пойдет речь.
Основные гипотезы
О биологической роли избыточной ДНК высказано не менее 15 гипотез. В первых из них (это версии Х.Кэллана, Х.Уайтхауза, Ч.Томаса, Р.Бриттена, Э.Дэвидсона, Г.П.Георгиева, Ф.Крика, П.Цукеркандла) фактически отрицалось наличие у такой ДНК неизвестных биологических функций. К излишку генома относили либо многократно повторенные гены (такие есть, но их очень немного), либо регуляторные зоны генетических единиц, подобные оперонам у бактерий. Однако уже в 60-70-х годах ХХ в. сопоставление известных фактов привело к заключению, что избыточная ДНК не выполняет ни кодирующих, ни регуляторных функций.
Гипотезу, не похожую ни на одну из упомянутых, предложил шведский биолог Эдстрем. По его мнению, избыточная ДНК представляет собой содержимое гипотетического “эволюционного котла”, в котором созревают новые структурные гены и новые регуляторные последовательности. Мысль эта не нова и имеет под собой некоторые основания. О них говорил еще в 20-е годы необыкновенно плодовитый на различные идеи русский генетик А.С.Серебровский. Действительно, новый ген не способен образоваться путем мутационной перестройки старого, если тот присутствует в единственном экземпляре. Если же существует избыток генетического материала в виде удвоенных (дуплицированных) генов, то один из них может и не функционировать, но в нем будут как бы тайно накапливаться мутации. Постепенно он преобразуется в новый ген, и когда в клетке создадутся условия для его работы, возникнет новый генный продукт, а следовательно, новый признак [2].
Итак, ни одна версия не была подтверждена. Но вот стало известно, что гены эукариот (некоторых архебактерий, а также эубактерий, которым несвойственна морфологическая эволюция, подобная эукариотической) состоят из экзонов (кодирующих участков) и интронов (некодирующих); что с каждого такого гена с помощью специального механизма - альтернативного сплайсинга - могут считываться несколько разных матричных РНК, а значит, синтезироваться несколько белков; что широко распространена транспозиция - перемещение мобильных элементов в геноме. Казалось бы, идея об избыточной ДНК как о сыром эволюционном материале с этими открытиями должна стать на твердую почву. Но ничего похожего не произошло. Дело в том, что львиная доля избыточной ДНК у большинства видов устроена по типу чередования (интерсперсии) некодирующих последовательностей - уникальных и повторяющихся. У человека такая часть генома занимает не менее 50%. У других видов содержание может отличаться, как и размеры чередующихся участков, причем какой-либо отчетливой закономерности не заметно. У морского ежа, африканской шпорцевой лягушки, человека и многих других видов короткие повторы в основном состоят примерно из 300 пар нуклеотидов, а уникальные последовательности из 1.5 тыс. У дрозофилы, рептилий, птиц намного длиннее и повторы (примерно 2000 пар нуклеотидов), и уникальные последовательности (5-10 тыс.). У двух видов плодовой мушки (D.melanogaster и D.virilis) типы повторов отличаются. А некодирующая часть генома шелкопряда больше похожа по параметрам чередования на геном человека, чем на геном D.melanogaster, хотя шелкопряд и дрозофила принадлежат к отряду чешуекрылых насекомых, а человек - к отряду приматов, высших млекопитающих. Это еще одна загадка, по сути, вероятно, тесно связанная с геномным парадоксом, и головоломка для исследователей, пытающихся найти функцию “лишней” ДНК исходя из традиционных представлений.
Скепсис в отношении разгадки геномного парадокса в начале 80-х годов достиг апогея. Может быть, именно потому появились гипотезы, согласно которым избыточная ДНК представляет собой “эгоистическую”, “паразитическую” или даже “мусорную” часть генома. Последний термин используется и сейчас, после расшифровки геномов человека, дрозофилы и других видов эукариот, не внесшей никакой ясности в объяснение парадокса. Между тем найти точные молекулярные характеристики “мусорной” ДНК в литературе просто невозможно. Мне представляется (в качестве рабочей гипотезы), что такая ДНК может иметь любую структуру, не содержащую последовательностей, которые способны блокировать репликацию ДНК. Впрочем, мусор в бытовом смысле - это то, что надлежит удалить, от чего необходимо избавиться. Но геномный “мусор” существует и, видимо, столько же, сколько существует вид. Более того, в гомологичных хромосомах “мусор” должен быть довольно сходным, чтобы не препятствовать их синапсису в мейозе, иначе гибридам придется расплачиваться стерильностью из-за различий в “мусорной” части генома. Это обстоятельство налагает сильные ограничения на мутационную дивергенцию “мусорной” ДНК.
Схема двух типов чередования нуклеотидных последовательностей разной степени повторяемости. Видно, что у шпорцевой лягушки, шелкопряда, мыши и человека некодирующие участки устроены сходным образом и повторы разной длины чередуются одинаково, только у дрозофилы чередование иное. 1 - уникальные последовательности, 2 - диспергированные повторяющиеся участки, n1 и n2 - короткие повторы; т.п.о. - тыс. пар оснований.Нельзя не упомянуть, пожалуй, еще одно соображение, касающееся роли избыточной ДНК, высказанное в 80-х годах английским ученым Т.Кавалье-Смитом [3]. Он обратил внимание на то, что некодирующие последовательности могут служить основой ядерного скелета, вокруг которого происходит сборка ядра. Это значит, что общее количество ДНК должно влиять на размеры ядра, клетки, скорость репликации и, в конечном счете, на размер тела и скорость развития организма. Таким образом, величина некодирующей части генома может оказаться под действием селективных факторов. Может быть, иногда это и так, особенно у некоторых растений. Но если сравнить, например, человека и мышь, геномы которых, как и доля некодирующих последовательностей в них, почти одинаковы, то без дальнейших обсуждений становится ясно, что гипотеза Кавалье-Смита и его последователей не может служить исчерпывающим объяснением геномного парадокса эукариот. Кроме того, она не объясняет сам механизм, который обеспечивал бы разное содержание ДНК.Диминуция хроматина - ключ к решению геномного парадокса?
Эфемерность многочисленных гипотез о биологической роли избыточной ДНК, скорее всего, обусловлена тем, что ни одна из них не опиралась на конкретный клеточный процесс. А ведь только на его основе можно найти хотя бы какие-то ответы на вопросы и определить направление дальнейших поисков. Полагаю, что такой процесс есть, более того, он известен очень давно. Это диминуция (уменьшение) хроматина, открытая выдающимся немецким биологом Т.Бовери в 1887 г. у аскариды: на самых ранних этапах развития из зародышевых клеток, которые потом станут соматическими, выбрасывается та или иная часть генетического материала. Надо сказать, что диминуция хроматина довольно редка, тем не менее сегодня она известна не только у аскарид, а еще у циклопов, миксин, сциарид.
Решающий вклад в открытие и детальное изучение диминуции хроматина у простейших внес Д.Прескотт [4, 5]. В начале 70-х годов прошлого века американский ученый выявил этот феномен у брюхоресничных инфузорий при созревании вегетативных (т.е. выполняющих соматические функции) ядер - макронуклеусов. У представителей родов Stylonуchia, Oxytricha, Euplotes эти ядра в процессе созревания теряют 96-98% нуклеотидных последовательностей, которые в генеративном ядре, микронуклеусе (аналоге клеток зародышевого пути у многоклеточных животных), сохраняются. Во время диминуции хромосомы макронуклеуса претерпевают фантастические изменения - попросту распадаются на отдельные гены, которые затем многократно копируются (амплифицируются), а некодирующие последовательности выбрасываются из ядра. Макронуклеус, оставшийся без “классических” хромосом, теряет способность делиться митозом и переходит к амитозу (прямому делению ядра перетяжкой). Митоз, который гарантирует получение дочерними клетками идентичного материнского набора единиц наследственной информации, перестает быть нужным, так как и при делении амитозом за счет большого избытка копий все гены сохраняются у клеточных потомков.
Столь колоссальная реорганизация генома у инфузорий казалась неким маргинальным событием, тупиком эволюции, не позволяющим делать эволюционные выводы о биологической роли избыточной ДНК.
С такой оценкой феномена геномной реорганизации у простейших мы с сотрудниками лаборатории в принципе не согласны. Поэтому продолжаем изучать диминуцию и ее роль, а объектами исследований нам служат типичные многоклеточные животные - веслоногие рачки циклопы [2, 6-9].
Мы не были первыми в изучении диминуции хроматина у циклопов. С.Берман это сделала еще в середине 60-х годов [10]. Немецкая исследовательница обратила внимание на то, что число хромосом у тех трех видов циклопов, с которыми она работала, было одинаковым как до диминуции, так и после нее. Конечно, за счет этого события размеры хромосом уменьшались в зависимости от доли потерянной ДНК. Берман предложила молекулярную модель диминуции: избыточная ДНК выводится из хромосом подобно тому, как профаг исключается из хромосом лизогенных бактерий, т.е. путем выпетливания и внутрихроматидной рекомбинации с образованием колец из ДНК. Кольца Берман и обнаружила при электронно-микроскопическом исследовании разрушенных клеток циклопов в стадии диминуции [11]. К сожалению, работы Берман прервались в 1984 г.
Схема строения самки веслоногого рачка циклопа. Яйцевые мешки, которые выходят из тела самки сразу после оплодотворения, содержат по 30-40 зародышей (на рисунке они похожи на шарики), в их соматических клетках на ранней стадии дробления и происходит диминуция хроматина. Сотрудник нашей лаборатории А.К.Гришанин обнаружил, что у рачка Cyclops kolensis в процессе диминуции из соматических клеток зародыша элиминируется рекордное для многоклеточных количество ДНК - 94% [7]. Это совсем ненамного меньше, чем у абсолютных рекордсменов - брюхоресничных инфузорий. Однако потеря огромной части ДНК не сказывается на числе хромосом: оно остается постоянным и равным 22 (столько же и в клетках зародышевого пути). Поразительно, как в одном организме циклопа могут одновременно существовать два типа клеток со столь разным содержанием ДНК!
Какие же наиболее важные выводы, касающиеся роли избыточной ДНК, можно сделать из исследования диминуции хромосом у циклопов?
Во-первых, для дифференцировки, гистогенеза и в целом построения всего тела C.kolensis достаточно лишь 6% хромосомной ДНК соматических клеток. Отсюда следует, что 94% их ДНК не содержат ни генов, ни регуляторных последовательностей, которые необходимы для индивидуального развития данного вида.
Во-вторых, диминуция хроматина количественно строго повторяется во время каждого цикла репродукции рачка (мы наблюдали это в течение более чем 10 лет). Такое повторение возможно лишь в том случае, если ДНК клеток зародышевого пути, не затронутая диминуцией, сохраняется столько времени, сколько существует вид. Иначе в соматических клетках никакого избавления от избыточной ДНК не происходило бы. Эти выводы - не результат косвенных умозаключений, они основаны на наблюдениях природного процесса.
Следовательно, ДНК, выводимая из хромосом во время диминуции, не может считаться “мусорной”. Более того, даже избыточна она только для соматических клеток, но не для клеток зародышевого пути. Несомненно, что и функции изымаемой ДНК надо искать именно в линии зародышевых клеток. Попытаемся это сделать, но прежде о самом процессе диминуции. Большинство биологов, видимо, знают лишь его конечный результат, т.е. что в соматических клетках ДНК меньше, чем в половых. В действительности диминуция - весьма сложный, многоэтапный процесс, который Дж.Шапиро справедливо отнес к наиболее ярким примерам природной генной инженерии [12]. Главное заключение, которое следует из феномена диминуции, очевидно: гены, участвующие в индивидуальном развитии, и обслуживающие их регуляторные участки ни в коем случае не должны быть ни потеряны, ни даже повреждены.
Разрезают и сшивают хромосомную ДНК ферменты диминуции. Они безошибочно осуществляют свои функции, поскольку места разрывов весьма точно помечены, т.е. участки ДНК заранее подготовлены. По масштабности и точности манипуляций, которые производят клеточные ферменты в ходе диминуции, подобная генно-инженерная операция пока недоступна исследователям, работающим в самой современной лаборатории.
Схема диминуции хроматина у циклопов. Сверху вниз: хромосома до начала диминуции (такова же и хромосома клеток зародышевого пути), во время диминуции, когда участки межгенной ДНК выпетливаются, и по окончании процесса: образовавшиеся на предыдущей стадии петли, вырезанные из хромосомы, превратились в кольца (масштаб не соблюден) [11]. S1, S2, S3 - структурные гены; перед ними в виде прямоугольников показаны регуляторные участки; мгДНК - межгенная ДНК, РХР - районы хромосомных разрывов.Что же произойдет, если ошибки все же возникнут? Летальные события: при повреждении гена эмбриональное развитие остановится тогда, когда наступит время функционирования этого гена. Примечательно, что диминуция происходит очень рано: у C.kolensis, например, на четвертом делении дробления, т.е. на 8-клеточной стадии, причем лишь в семи клетках, а восьмая, дающая начало зародышевой линии, остается с исходными хромосомами. (У рачка C.strenuus strenuus Гришанин наблюдал диминуцию, проходящую в два этапа - на пятом делении удаляется 40% ДНК, на следующем еще 35%. Есть и другие виды с подобной двухэтапной диминуцией.)Второй вариант ошибочного разрезания - хромосомные перестройки. Это тоже летальные события. Однако, по нашим данным, число спонтанных хромосомных перестроек в раннем развитии циклопов поразительно мало [8]: примерно в 100 раз меньше, чем у такого классического объекта для тестирования хромосомного мутагенеза, как лимфоциты человека. Этот факт свидетельствует о наличии мощной защитной системы против мутаций, действующей по крайней мере тогда, когда в клетке высока активность ферментов диминуции. Они должны атаковать только предназначенные для этого районы ДНК, иначе могут появиться поврежденные участки, которые окажутся чувствительными к таким ферментам и будут ими разрезаны. Следовательно, подобная возможность, грозящая катастрофическими последствиями, должна быть сведена до минимума.
А если бы диминуции подверглась та единственная клетка, которая предназначена для развития зародышевой линии? Тогда опасность таилась бы не для особи, а для вида, поскольку исключительно высока, почти неизбежна, стерильность индивида, у которого произошла подобная незапрограммированная потеря генетического материала.
Что находится в гранулах?
У C.kolensis вырезанные из хромосом участки ДНК собираются в гранулы, окруженные уникальной - плотной и без пор - мембраной [9]. Их содержимое, как мы полагаем, и есть избыточная ДНК, которая создает парадокс размера генома у эукариот. Клеточные механизмы отличают эту ДНК от той, что содержится в хромосомах клеток зародышевого пути и не должна быть потеряна, пока существует данный вид.
Итак, мы с коллегами убедились, что у циклопов во время раннего эмбриогенеза из соматических клеток удаляется и сосредоточивается в специфических гранулах бoльшая часть ДНК. Но связаны ли с ней какие-либо функции, мы не знали, их предстояло изучить. Мы начали с исследования содержимого гранул (т.е. элиминируемой ДНК) методами геномики и протеомики [13]. В этой работе принимала участие группа новосибирских ученых из Института цитологии и генетики СО РАН, которую возглавлял И.Ф.Жимулев. Приятно назвать имена наших коллег: Н.Б.Рубцов, Т.В.Бойкова, С.Н.Белякин, Т.Карамышева. Существенную помощь нам оказал также английский генетик М.Ашбернер из Кэмбриджа.
Микрофотография хромосом Cyclops kolensis в клетке зародышевого пути (внизу слева) и в анафазе соматических клеток (справа) во время диминуционного деления. На врезке показана гранула с элиминированной ДНК. Видно, что гранула окружена плотной однослойной мембраной, лишенной пор.Объектом совместных исследований стал все тот же рачок C.кolensis. Гранулы с элиминированной ДНК, образовавшиеся во время четвертого деления дробления, были извлечены из клеток с помощью микроманипулятора, а полученные затем индивидуальные фрагменты ДНК клонированы. Нуклеотидная последовательность, установленная в каждом из них, подверглась компьютерному анализу, и в результате выяснилось, что генных участков в них нет. Иного мы и не ожидали. Вся та ДНК, которую мы изучали, по старой терминологии должна быть отнесена к “мусорной”.

Удаление гранул элиминированного хроматина
во время пятого деления дробления при помощи микроманипулятора.Однако по результатам анализа индивидуальных фрагментов и их сравнения между собой вырисовывалась другая картина, далеко не та, что соответствует концепции молекулярного “мусора” [14]. Набор фрагментов представлял собой вовсе не “свалку” случайных нуклеотидных последовательностей.
В индивидуальных фрагментах, длина которых составляла 400-500 пар нуклеотидов, содержались относительно небольшие повторы - до 34 нуклеотидных пар (повторы менее семи пар не учитывались, поскольку они могли возникнуть случайно). Многие повторы характеризовались высокой степенью гомологии, а некоторые оказались абсолютно идентичными.
Мы не исключали, что сходные по последовательности участки одного клонированного фрагмента найдутся и в других. Всего их было 52, и вместе они составляли примерно 32 тыс. нуклеотидных пар. Оригинальный поиск копий среди такого множества выполнил Дегтярев с коллегами [14].
Поиски дали замечательный результат: в исходных фрагментах появились новые, более длинные повторы, содержащие до 296 пар нуклеотидов. Такая метаморфоза могла произойти лишь в том случае, если эти копии последовательностей состояли из более мелких повторов (субповторов), идентифицированных нами в индивидуальных фрагментах и разбросанных по геному. Следовательно, структура длинных повторов мозаична. Различия же в их нуклеотидной последовательности оказались весьма незначительными. Более того, один из самых протяженных субповторов - в 57 пар нуклеотидов - присутствовал в виде точных копий везде, где бы его ни обнаруживали.
Мозаичность структуры отдельных отрезков элиминируемой ДНК и высокая степень гомологии повторяющихся последовательностей наводят на мысль, что к качественным изменениям, которые должен непрерывно генерировать всесильный мутационный процесс, “мусорная” часть генома удивительно устойчива.
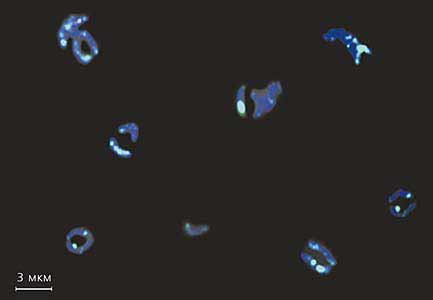
Гибридизация in situ клонов, полученных из элиминированной ДНК циклопа, с хромосомами, еще не подвергшимися диминуции. Светлоокрашенные участки - места, где расположена элиминированная ДНК.Каким образом могут возникать последовательности с высокой степенью гомологии? Их образование относят на счет концертной эволюции и рассматривают обычно две причины, ведущие к ней. Первая - недавнее происхождение повторов от общей предковой последовательности: просто не хватило времени на то, чтобы в немногих поколениях накопились мутации и в результате утратилось сходство. Вторая причина - рекомбинации: если гомологичные участки расположены в ядре клетки так, что между ними возможны эти процессы, при определенных обстоятельствах вновь возникающие мутации будут уничтожаться, т.е. гомологи сохранятся [15].
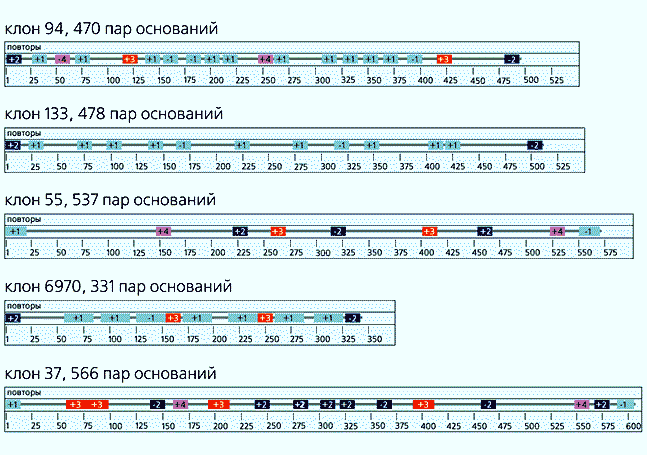
Пример повторяющихся последовательностей внутри каждого из пяти приведенных фрагментов элиминированной ДНК Cyclops kolensis. Гомологичные повторы в конкретном фрагменте показаны одним цветом, в разных фрагментах одинаковая окраска не означает гомологии. Цифры внутри блоков - номера повторов, знаком перед цифрой отмечена их ориентация - прямая (+) или обратная (–). Числа на шкале отражают количество пар оснований.
Схема расположения повторов в разных фрагментах элиминированной ДНК Cyclops kolensis. Повторяющиеся последовательности принадлежат 24 семействам, их номера указаны числами, знаком “+” отмечена прямая ориентация повтора, знаком “–” - обратная. В первом столбце - номера клонов. Шкала градуирована по количеству пар нуклеотидов. Праймерные последовательности, необходимые для клонирования, обозначены цифрой 1.Какая из этих причин способна привести к высокой степени гомологии повторов в избыточной ДНК при том, что любому виду свойственны постоянно протекающие локальные геномные перестройки? Такие реорганизации генома у видов с диминуцией в состоянии изменить саму структуру районов, где обязан происходить разрыв хромосом. У C.kolensis, например, подобных районов должно быть не менее 10 тыс., как показывают наши расчеты. Геномные перестройки могут привести к таким изменениям сигнальных участков для диминуции, что их перестанут узнавать ферменты. А за этим последует гибель организма на ранней стадии развития. Возможно, подобные перестройки создадут область разрыва совсем не в том месте, где она должна располагаться в норме. Тогда ферменты диминуции произведут ошибочные операции, что вызовет в итоге гибель всех потомков мутантной клетки. Поэтому нам представляется более предпочтительным механизм концертной эволюции генома циклопов через консервацию повторов за счет рекомбинаций.Наши данные по молекулярной структуре элиминируемой ДНК, полученные, кстати, впервые, все же следует считать предварительными. Тем не менее они довольно ясно указывают на высокую степень упорядоченности организации той части генома циклопа, которая изымается при диминуции и которая могла бы считаться образцом молекулярного “мусора”.
Каково же значение избыточной ДНК? Наличие этой ДНК в клетках зародышевого пути создает уникальный геномный портрет вида и, следовательно, может служить надежным механизмом его генетической изоляции [6, 16]. Если бы такой портрет не сохранялся в ряду поколений, синапсис гомологичных хромосом в мейозе нарушался бы, и потомство становилось анеуплоидным (т.е. с некратным гаплоидному числом хромосом), без каких-либо шансов на выживание. * * *
Осталось коснуться проблемы, которая связана с колебаниями размеров генома в эволюции. Благодаря чему может быстро уменьшаться размер генома? Мы полагаем, что объяснением этого наиболее трудного аспекта эволюционной проблемы служит диминуция хроматина [17]. Если изъятию подвергнется ДНК клетки-родоначальницы зародышевого пути и процесс не будет сопровождаться летальным событием, поскольку коснется лишь некодирующей ДНК, то все гаметы данного организма окажутся обладательницами нового - редуцированного - генома. Его получат сразу многие десятки особей, а это может быстро создать изолированную группу.
И еще об одном нельзя не сказать. Неужели циклопы, чей онтогенез сопровождается диминуцией хроматина, так сильно отличаются от других животных, у которых нет такого же способа физически исключить избыточную для соматических клеток часть генома? Едва ли это можно утверждать. У других видов существуют иные способы инактивировать, т.е. в конечном счете заставить замолчать, эту ДНК. Известно, что у человека около 50% генома вообще не участвует в экспрессии. Целые районы генома оказываются недоступными для транскрипции чаще всего из-за сверхкомпактной укладки участков хромосом. Способ другой, но результат тот же - изоляция участков генома.
Тут уместно вспомнить слова знаменитого П.Тейяра де Шардена из его книги “Феномен человека” (1965): “Природная аномалия - это всегда лишь преувеличение до ощутимости какого-либо свойства, всюду распространенного в неосязаемом виде. Какой-либо феномен, точно установленный хотя бы в одном месте, в силу фундаментального единства мира имеет повсеместные корни и всеобщее значение”. Разве диминуция хроматина не относится к числу твердо установленных феноменов? Ведь она протекает на глазах исследователей!
Современный этап в изучении диминуции еще только начинается, хотя известно о ней было давно, а 30 лет назад уже делались предположения о роли избыточной ДНК [2]. Cейчас стали просматриваться эволюционные перспективы, которые в данной статье были перечислены далеко не полностью.
Работы поддержаны Российским фондом фундаментальных исследований (проекты 01-01-00227 и 03-03-48133)
и Государственным контрактом (№10002-251/П-25/155-154/200404-071).
Литература1. Mirsky A.E., Ris H. // Gen. Physiol. 1951. V.34. P.451-462.
2. Акифьев А.П. Молчащая ДНК и ее роль в эволюции // Природа. 1974. №9. С.49-54.
3. Cavalier-Smith T. // Ann. Rev. Biophys. Bioeugin. 1982. V.11. P.273-302.
4. Prescott D. M., Nuryi K.G., Bostook C.J. // Nature. 1973. V.376. P.597-600.
5. Prescott D.M. // Current opinion in genet. and Devel. 1997. V.7. P.807-813.
6. Акифьев А.П., Гришанин А.К., Дегтярев С.В. // Генетика. 1998. Т.34. С.709-718.
7. Гришанин А.К., Худолий Г.А., Шайхаев Г.О. и др. // Генетика. 1996. Т.32. С.492-499.
8. Гришанин А.К., Дегтярев С.В., Акифьев А.П. // Генетика. 2002. Т.38. С.468-472.
9. Гришанин А.К. // Онтогенез. 1995. Т.26. С.188-195.
10. Beermann S. // Chromosoma. 1977. V60. P.297-344.
11. Beermann S. // Chromosoma. 1984. V.89. P.321-328.
12. Shapiro J.A. // Genetica. V.86. P.96-111.
13. Дегтярев С.В., Гришанин А.К., Белякин С.Н. и др. // Докл. Акад. наук. 2002. Т.384. С.255-258.
14. Degtyarev S., Boykova T., Grishanin A. et al. // Genome Res. (in press.)
15. Murti J.R., Bumbulis M., Schimentic C. // Genetics. 1994. V.137. P.837-843.
16. Акифьев А.П., Гришанин А.К., Дегтярев С.В. // Генетика. 2002. Т.38. С.595-606.
17. Gregory T.R. // Trends in genetics. 2003. V.9. P.485-488.