№10, 2002 г.
№10, 2002 г.
№10, 2002 г.
№10, 2002 г.
© М.В.Деренко, Б.А.Малярчук
ГЕНЕТИЧЕСКАЯ ИСТОРИЯ
КОРЕННОГО НАСЕЛЕНИЯ
СЕВЕРНОЙ АЗИИМ.В.Деренко, Б.А.Малярчук
Мирослава Васильевна Деренко, кандидат биологических наук,
ведущий научный сотрудник лаборатории генетики.
Борис Аркадьевич Малярчук, кандидат биологических наук,
ведущий научный сотрудник той же лаборатории.История коренных народов Северной Азии связана, как теперь установлено, и с развитием аборигенных неолитических племен, и с бурными этногенетическими событиями на юге Сибири, и с великими переселениями народов, когда носители культур бронзы и железа проникали на Крайний Север. Значительную роль играло также внедрение отдельных групп южного происхождения в среду северных этносов. И хотя этнические черты более или менее отчетливо начинают проступать в бронзовом веке, генетическая история народов уходит своими корнями в гораздо более древние эпохи.
Согласно археологическим и палеоантропологическим данным, Северную Азию человек современного типа начал заселять с верхнего палеолита. Однако число известных археологических памятников с этих территорий невелико, и все они сравнительно однородны, за исключением двух стоянок — Мальты и Бурети на Ангаре и ее притоке Белой. Археологи отмечают, что там традиции в пещерной живописи, обработке камня и т.д. напоминают европейские. Тем не менее в североазиатском верхнем палеолите преобладали архаичные формы каменного инвентаря. Палеоантропологические находки этого времени довольно редки и фрагментарны, однако в целом они свидетельствуют о доминировании монголоидных признаков у древнего населения Северной Азии. Между тем ряд фактов указывает на то, что степная часть Центральной Азии — от Алтая, Хакасии, Тувы и до Внутренней Монголии — издревле была ареной взаимодействия групп монголоидного и европеоидного происхождения и, более того, представляла собой зону различных очагов европеоидного расообразования [1]. Популяции европеоидного и смешанного происхождения уже широко расселились на указанных территориях в эпоху неолита, бронзы и раннего железа. Однако культурная преемственность археологических памятников уводит генезис неолитического населения Монголии и Алтае-Саянского нагорья к более раннему времени [1, 2].
На формирование антропологического облика современного населения Северной Азии в разное время заметное влияние оказывали отдельные группы европеоидов: в Западной Сибири это были восточноевропейские группы, а на юге Сибири, вероятно, южные европеоиды из Казахстана и Средней Азии.
Комплекс антропологических черт монголоидной расы у североазиатского населения тоже неоднороден. Связано это с тем, что в эпоху позднего палеолита в Азии формировался восточный очаг расообразования, а дальнейшая дифференциация монголоидов происходила в нескольких очагах. Один из них располагался в Центральной Азии и Забайкалье и рассматривался антропологами в качестве прародины сибирской общности народов. Последовательная дифференциация населения региона, начавшаяся в неолите, привела к возникновению многих расовых типов, которые и наблюдаются у современных этносов Сибири. Но нельзя исключать, что на формирование их антропологического облика влияли тихоокеанские монголоиды, комплекс признаков которых довольно четко проявляется в некоторых группах жителей Чукотки, Камчатки, Охотоморья и Приморья.
Итак, коренным народам Северной Азии присуще высокое антропологическое разнообразие, их этнолингвистическая история сложна, а область расселения, которое сопровождалось интенсивным межэтническим взаимодействием, обширна. Все это должно быть зафиксировано и в генетической истории североазиатских этносов.
Генетические маркеры народов
Попробуем проследить генетическую историю и используем для этого анализ изменчивости митохондриальной ДНК (мтДНК) и нерекомбинирующих областей Y-хромосомы.
Данные об изменчивости мтДНК (скорость накопления мутаций в ней в 5—10 раз превышает таковую в ядерных генах) служат для точной идентификации женских линий и детального описания генетических компонентов в смешанных популяциях. К настоящему времени установлено, что митохондриальные генофонды популяций человека представлены несколькими наборами групп из определенных типов (линий) ДНК, причем каждая из них ведет свое происхождение от единственной основательницы. Для большинства групп типов мтДНК установлена региональная специфичность. Так, в генофондах народов Европы и Западной Азии (т.е. европеоидов) присутствуют главным образом 10 основных групп мтДНК (HV*, H, V, J, T, U, K, I, W, X), в генофондах монголоидных популяций Азии — тоже 10, но других групп (A, B, C, D, E, F, G, Y, Z, M*), а у негроидов — мтДНК макрогруппы L [3—5]. Заметим, что коренное население Америки характеризуется редуцированным набором групп — A, B, C, D и X. Чтобы определить, откуда и когда происходили миграции, которые привели к современному генофонду анализируемой популяции, необходимо учитывать, что при его формировании была перенесена часть генетического разнообразия из популяций-основательниц. Кроме того, нужно располагать результатами филогеографического анализа мтДНК.
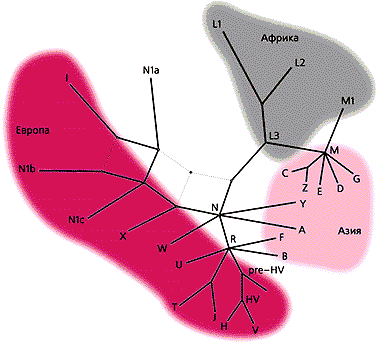
Эволюционное дерево митохондриальной ДНК человека.
Мужской “аналог” мтДНК — Y-хромосома, наследуемая по отцовской линии. Наиболее информативен для целей этнической генетики нерекомбинирующий участок Y-хромосомы (его длина превышает 60 млн пар нуклеотидов), в котором уже идентифицировано более 250 полиморфных локусов. В их числе и медленно эволюционирующие диаллельные маркеры, и быстро эволюционирующие микро- и минисателлитные локусы. Следовательно, мутации, накапливающиеся в мужских линиях в процессе эволюции, тоже служат важным источником информации об основных этапах дифференциации этнорасовых групп (популяций) и о происхождении конкретных индивидуумов. К настоящему времени установлено, что распределение маркеров Y-хромосомы специфично для разных популяций мира. Например, только коренному населению Америки свойствен уникальный вариант этой хромосомы с мутацией в локусе DYS199, наличие которой свидетельствует о едином происхождении большинства мужских линий. Более того, удалось выяснить, что предковыми вариантами “американского” типа мужской хромосомы были палеоевропеоидные линии. Именно на их основе сформировались и некоторые варианты Y-хромосомы, распространенные в современных азиатских и европейских популяциях.
Что касается коренных народов Сибири, то большинство работ, посвященных анализу полиморфизма мтДНК или Y-хромосомы, связаны лишь с проблемой происхождения аборигенного населения Америки. И только в последние годы изучение эволюционной истории конкретных этнических групп и жителей отдельных регионов Северной Азии стало самостоятельной научной задачей [6—10].
Сходства и отличия по материнской линии
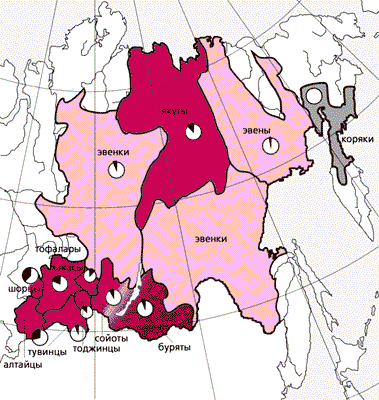
Мы исследовали структуру и разнообразие генофондов 12 этнических групп коренного североазиатского населения. Суммарную выборку составили 936 человек из различных лингвистических групп алтайской и палеоазиатской языковых семей. Первую семью образовали три группы: тюркская (алтайцы, хакасы, шорцы, тувинцы, восточные тувинцы, или тоджинцы, тофалары, сойоты и якуты), монгольская (буряты) и тунгусо-манчжурская (эвенки и эвены). Представителями палеоазиатской языковой семьи были коряки.
Ареалы североазиатских народов и диаграммы, отражающие соотношение монголоидных (белый фрагмент) и европеоидных линий мтДНК в каждой этнической группе. Генофонды этих народов, судя по мтДНК, содержат обе линии в разной пропорции, а по лингвистической принадлежности составляют две языковые семьи — алтайскую и палеоазиатскую, причем к последней принадлежат только коряки. Первую же семью образуют три языковые группы: тюркская (алтайцы, хакасы, шорцы, тувинцы, восточные тувинцы, или тоджинцы, тофалары, сойоты и якуты), монгольская (буряты) и тунгусо-манчжурская (эвенки и эвены).Митохондриальные генофонды сибирских популяций оказались весьма разнообразными. Наряду с типично монголоидными группами мтДНК (M*, M7, M8, M9, M10, C, D, G, Z, A, B, F, N9a, Y), частота встречаемости которых меняется от 64.3% у шорцев до 92.4% у эвенков, присутствуют и европеоидные линии (H, HV, I, J, K, T, U, N1a, X). Примечательно, что частота этих линий мтДНК увеличивается в западном направлении и достигает максимума (35.7%) в популяциях шорцев и алтайцев. Наличие множества генетических вариантов обусловлено в первую очередь разнообразием комбинаций групп мтДНК, но преобладает обычно одна из них. Так, в генофондах бурят и сойотов доминирует группа D, у тувинцев, тоджинцев и тофаларов — группа C (у последних ее частота наивысшая в Азии — 62%), у шорцев — типы мтДНК из группы F (42.9%), с высокой частотой (22%) обнаруженные также у хакасов. Интересно, что ранее эти типы отмечались (с частотами до 32%) только у южных китайцев и вьетнамцев.Уникальность генофондов сибирских популяций — присутствие нескольких подгрупп мтДНК из группы G. У коряков и эвенов с высокими частотами встречается подгруппа G1, у бурят — G2, а у восточносаянских народов — G3 и G4. Подобная разветвленность мтДНК группы G свидетельствует в пользу ее значительной древности в генофондах сибирских народов. Возраст подгруппы G2 составляет не менее 27 тыс. лет, т.е. произошла она в позднем палеолите, а значит, группа G в целом — еще раньше. Мы обнаружили и два варианта группы Z. Один из них (Z1), самый древний, найден у населения Южной Сибири, второй (Z2), более молодой, — у жителей ее северо-восточной части, причем у эвенов частота Z2 достигает 30%. Интересно, что Z2-типы довольно широко распространены в генофондах народов Северной Европы, в том числе саамов, имеющих в облике черты монголоидной расы. У них кроме Z2 обнаружена еще и специфическая подгруппа D5 [11]. Важно отметить, что и Z2-, и D5-типы мтДНК — компоненты генофондов алтае-саянских народов. А это наводит на мысль об их участии в этногенезе саамов.
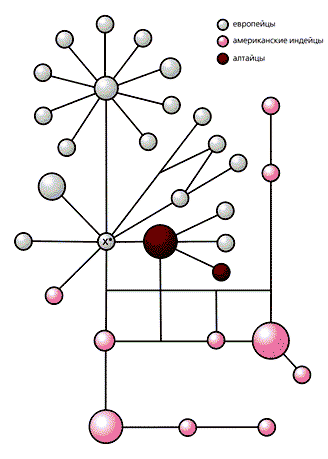
МтДНК группы B, входящие в состав монголоидного компонента сибирских популяций, раньше были выявлены у народов Центральной и Восточной Азии, а также у индейцев Америки. В южносибирских популяциях ее обнаружил Р.И.Сукерник с коллегами в 1996 г. В настоящее время установлено, что ареал носителей группы B в Южной Сибири охватывает Алтае-Саянский и Байкальский регионы, вплоть до Якутии, а максимальная частота этой мтДНК, зарегистрированная нами у западных тувинцев, составляет около 13%. Мало того, у южносибирских народов встречаются две разновидности группы B — B1 и B2, которые широко распространены и в более южных, центральноазиатских, популяциях. О давних генетических связях между населением Южной Сибири и Центральной и (или) Восточной Азии свидетельствует еще один факт: “неканонические” группы мтДНК из макрогруппы M (M7, M8, M9, M10, M*), характерные преимущественно для этнических общностей Китая, мы обнаружили у южносибирских народов. Между тем в Южной и Восточной Сибири мы выявили митохондриальные линии, не известные прежде у других азиатских популяций. В первую очередь это группы N1a, I и X, которые принадлежат к одному из наиболее древних “узлов” филогенетического дерева мтДНК человека. Эти группы в полном составе присутствуют в генофонде алтайцев. Заметим, что наличие у них X-мтДНК проясняет историю происхождения коренного населения Америки. Ранее было показано, что группа X распределена мозаично: у населения Европы, Кавказа, Западной Азии (с невысокими частотами), но и у североамериканских индейцев якима, навахо, нуу-чах-нулс, оджибва (до 30%). Поскольку X-мтДНК найдена также в костях из захоронений доколумбовой эпохи, эта группа не могла быть привнесена в генофонд индейцев современными европеоидами. Детальный филогенетический анализ нуклеотидных последовательностей свидетельствует о едином происхождении европейских и американских X-мтДНК, а время их расхождения оценивается в 12—36 тыс. лет [12]. Таким образом, группа X вместе с A, B, C и D относится к числу пяти линий-основательниц, сформировавших генофонды коренного населения Америки. Прежде группу Х не находили ни в Центральной, ни в Восточной Азии, включая Сибирь, и потому считалось, что в заселении Америки участвовали древние европеоиды, не оставившие генетических следов своего пребывания в Азии. Но алтайские варианты X-мтДНК, отличные как от европейских, так и от американских, занимают промежуточное положение между ними в филогенетических построениях [13]. Тем самым опровергается возможность появления этих вариантов за счет современной европеоидной примеси. Напротив, наши данные свидетельствуют, что в первичном заселении Америки участвовали выходцы из Южной Сибири, антропологический состав которых уже тогда, в эпоху верхнего палеолита, был разнородным.
В целом, судя по изменчивости мтДНК у населения Северной Азии, его генофонды формировались на смешанной генетической основе. Ее составили не только разнообразные монголоидные линии, но и европеоидный компонент, который выражен в разной степени в большинстве исследованных популяций.
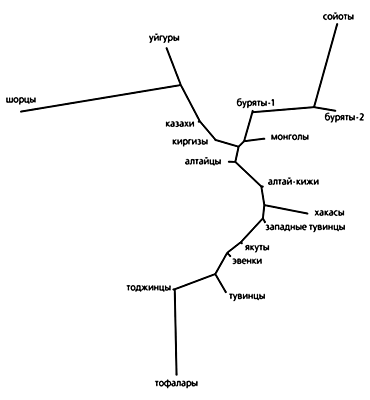
Гетерогенность генофондов свойственна и коренному населению Южной Сибири, за исключением тофаларов — географически изолированной группы восточносаянских тюрков. Низкое генетическое разнообразие тофаларов может быть обусловлено их малочисленностью, обособленностью и эффектом инбридинга. Известно, что население пос. Алыгджер, исследованного нами, сформировалось в основном из двух небольших довольно изолированных инбредных родоплеменных групп: Чогда и Кара-Чогда. Очень высокая частота (62%) типов мтДНК группы C у их представителей свидетельствует, что генофонд основан малым числом женских линий. Хотя генетически тофалары обособлены от других сибирских народов, в филогенетическом дереве они соседствуют с географически ближайшими к ним тувинцами и тоджинцами.
Филогенетическая сеть линий мтДНК группы X (предковый тип обозначен звездочкой), отражающая связь между европейцами, американскими индейцами и алтайцами. Алтайские X-линии, как видно, занимают промежуточное положение между европейскими и североамериканскими, что свидетельствует о заселении Америки из Центральной (или Южной) Сибири. Размеры кружков пропорциональны количеству индивидуумов, имеющих определенный тип мтДНК. Подобная дифференциация восточносаянских этнических групп отмечалась и ранее (по белковому полиморфизму). Ю.Г. Рычков с коллегами полагали, что современная популяционно-генетическая картина Восточных Саян сложилась из трех генетически разобщенных групп, следы этого смешения сохраняются в генофондах тофаларов, тоджинцев и тувинцев [14]. Впоследствии исходная гетерогенность усилилась за счет случайных генных миграций, происходивших от Тувы через Тоджу к Тофаларии. Это подтверждается и другими данными. Так, антропологический тип меняется в том же направлении: тувинцы принадлежат к центральноазиатскому типу, тоджинцы — к смешанному, тофалары — к катангскому варианту байкальского типа. Однако лингвистическое замещение, подобное смене самодийского языка в Тофаларии на тюркский, который проник с юга, из-за хребта Восточного Саяна, по всей видимости, не сопровождалось сменой населения и нарушением генетической преемственности. Например, тюркоязычные сойоты генетически сходны с монголами и монголоязычными бурятами, а западные эвенки, относящиеся к тунгусо-манчжурской группе языков, в генетическом отношении близки к их восточным соседям — якутам. Возможно, сходство генофондов западных эвенков, тувинцев и тоджинцев обусловлено участием в их этногенезе тюркоязычного населения из Центральной Азии. Об этом же свидетельствует наличие в генофонде западных эвенков (и якутов) мтДНК группы B, которая наиболее распространена среди центральноазиатских и южносибирских народов, но отсутствует у тунгусо-манчжурских по языку эвенов, живущих на северо-востоке Азии. Некоторые несоответствия в филогенетическом положении бурят, тувинцев, якутов и западных эвенков прослежены и по антропологическим типам. Первые три народа (и частично тофалары) относятся к центральноазиатскому антропологическому типу, в то время как эвенки — к байкальскому. Между тем высокое генетическое сходство западных эвенков и якутов может быть результатом формирования их генофондов на основе почти одних и тех же материнских линий мтДНК. Ведь известно, что якуты по мере их расселения в Сибири ассимилировали многих эвенков.
Филогенетическое дерево, построенное на основе изменчивости мтДНК и иллюстрирующее эволюционные взаимоотношения этнических групп Сибири, Средней (уйгуры, казахи, киргизы) и Центральной Азии (монголы). Этнические связи по Y-хромосоме
Генетическая история населения Южной и Восточной Сибири еще ярче, чем в изменчивости мтДНК, отражена в разнообразии мужской половой хромосомы. Мы исследовали вариабельность 15 диаллельных локусов нерекомбинирующей части Y-хромосомы, которые характеризуют ее основные группы у населения Евразии. Группы Y-хромосомы (традиционно их обозначают HG — от англ. haplogroup), как и мтДНК, — это отдельные линии ДНК, идущие от общего предка, но отличающиеся друг от друга мутациями. Из-за низкой мутационной скорости диаллельных локусов по Y-группам удается исследовать самые ранние этапы истории человечества.
В обобщенном генофонде коренного населения Северной Азии мы выявили 12 линий Y-хромосомы, причем почти у всех сибирских аборигенов обнаружены группы HG1 и HG3. С максимальными частотами они встречаются у южных алтайцев (алтай-кижи и телеутов) и шорцев, с минимальными — у бурят. Известно, что T-аллель локуса 92R7, полиморфизмом которого маркируется группа HG1, широко распространен среди населения Евразии, а также индейцев Америки и представляет собой древнюю палеоевропеоидную ветвь, связанную своим происхождением с народами Центральной Евразии [15].HG1 обнаружена у населения Индии (частота 21.7%), Европы (средняя частота 37%, а у ирландцев она достигает 98.5%), Западной Сибири (у кетов 83.3%, у селькупов 76.2%), а также у аборигенов Америки (средняя частота 35.2%) [15—17].
Исследователи полагают, что подобная распространенность 92R7-T-типов Y-хромосомы может быть обусловлена как древними миграциями населения из Центральной Евразии в Европу и Америку, так и относительно недавним (примерно 7.5 тыс. лет назад) проникновением представителей курганной культуры из Центральной Азии на Индийский субконтинент, в Европу и Сибирь.
Что касается группы HG3, идентифицируемой полиморфизмом гена SRY1532, то она практически отсутствует у этносов Африки, Восточной Азии и Америки, но обнаружена с высокими частотами в Центральной и Восточной Европе (до 50% у русских, поляков и словаков), Юго-Западной Азии (до 32% в популяциях Индии, Пакистана и Ирана) и на Алтае (с частотой 55%) [15—17].
Рассмотрим теперь распределение еще нескольких групп Y-хромосомы, распространенных у населения Северной Азии. Группа HG10 преобладает, как установлено в наших исследованиях, в генофондах бурятов и сойотов, живущих в Байкальском регионе, а также эвенов и коряков из Северного Охотоморья. HG10, наряду с группой HG1, рассматривается в качестве одного из главных “основателей” мужского генофонда аборигенов Америки. Здесь ее появление, полагают Т.М.Карафет с коллегами, связано с миграциями из Восточной Сибири, включая Приамурье. Именно в этом регионе обнаружены варианты группы HG10, наиболее близкие тем, что наблюдаются, например, у индейцев навахо.
Таким образом, в настоящее время с достаточной долей уверенности можно считать, что палеопопуляции Южной и Восточной Сибири были источником нескольких (по меньшей мере — двух) миграций в Америку. В ходе одной из миграций — алтае-саянской — могли попасть, среди прочих типов Y-хромосомы и мтДНК, палеоевропеоидные мужские линии группы HG1, на основе которой сформировался чисто “американский” тип Y-хромосомы — группа HG18. В период другой волны расселения, шедшего из Байкальского региона, по всей видимости, распространилась группа HG10.
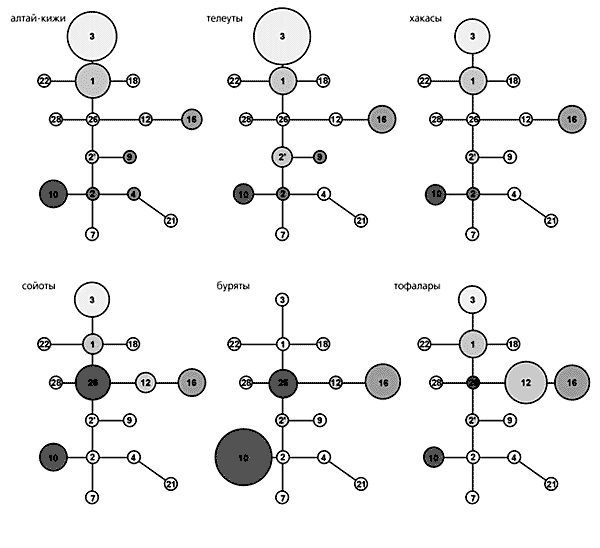 Филогенетические схемы, построенные по изменчивости Y-хромосомы. Видно, что проанализированные 15 гаплогрупп (обозначены числами) встречаются в генофондах сибирских народов с разной частотой (она пропорциональна размеру кружка).Для современного населения Сибири характерна одна из наиболее частых в Северной Евразии (от Северо-Восточной Азии до Восточной Европы) группа HG16. В генофондах этнических общностей Южной и Восточной Сибири ее частота меняется от 2% у шорцев до 55.6% у якутов; у коряков и эвенов ее частота составляет около 30%. Примечательно, что в генофондах этносов Алтае-Саянского региона присутствует также группа HG12 (предковая по отношению к HG16), максимальная частота которой — до 35% — зарегистрирована нами у шорцев и тофаларов. Необходимо отметить, что группы HG12 и HG16 нельзя однозначно отнести к числу монголоидных или европеоидных, да и их генетический вклад в происхождение народов Северной Евразии трактуется по-разному, равно как и само возникновение групп. Тем не менее обнаруженная нами высокая частота встречаемости группы HG12 и наличие группы HG16 в алтае-саянских популяциях могут указывать на их сибирские корни. Не исключено, что в генофонды народов Восточной Европы эти группы привнесены гуннами, чьи миграции документированы исторически.
Филогенетические схемы, построенные по изменчивости Y-хромосомы. Видно, что проанализированные 15 гаплогрупп (обозначены числами) встречаются в генофондах сибирских народов с разной частотой (она пропорциональна размеру кружка).Для современного населения Сибири характерна одна из наиболее частых в Северной Евразии (от Северо-Восточной Азии до Восточной Европы) группа HG16. В генофондах этнических общностей Южной и Восточной Сибири ее частота меняется от 2% у шорцев до 55.6% у якутов; у коряков и эвенов ее частота составляет около 30%. Примечательно, что в генофондах этносов Алтае-Саянского региона присутствует также группа HG12 (предковая по отношению к HG16), максимальная частота которой — до 35% — зарегистрирована нами у шорцев и тофаларов. Необходимо отметить, что группы HG12 и HG16 нельзя однозначно отнести к числу монголоидных или европеоидных, да и их генетический вклад в происхождение народов Северной Евразии трактуется по-разному, равно как и само возникновение групп. Тем не менее обнаруженная нами высокая частота встречаемости группы HG12 и наличие группы HG16 в алтае-саянских популяциях могут указывать на их сибирские корни. Не исключено, что в генофонды народов Восточной Европы эти группы привнесены гуннами, чьи миграции документированы исторически.* * *
Итак, благодаря молекулярно-генетическим исследованиям к настоящему времени детально охарактеризованы генофонды многих этнорасовых групп. Казалось бы, по распределению маркеров мужских и женских линий ДНК в современных популяциях человека, можно зафиксировать лишь последний кадр их истории. Но филогеографический анализ молекулярно-генетических данных позволяет проследить процесс формирования генофондов в ретроспективе. А значит, и познать историю народов мира и их общностей.
Работа выполнена при поддержке Российского фонда фундаментальных исследований. Проект 99-06-80430.
Литература
1. Алексеев В.П. Историческая антропология и антропогенез. М., 1989.
2. Алексеев В.П., Гохман И.И. Антропология азиатской части СССР. М., 1984.
3. Wallace D.C. // Am. J. Hum. Genet. 1995. V.57. P.201—223.
4. Macaulay V., Richards M., Hickey E. et al. // Am. J. Hum. Genet. 1999. V.64. P.232—249.
5. Yao Y.-G., Kong Q.-P., Bandelt H.-J. et al. // Am. J. Hum. Genet. 2002. V.70. P.635—651.
6. Schurr T.G., Sukernik R.I., Starikovskaya Y.B., Wallace D.C. // Am. J. Phys. Anthropol. 1999. V.108. P.1—39.
7. Derenko M.V., Malyarchuk B.A., Dambueva I.K. et al. // Hum. Biol. 2000. V.72. P.945—973.
8. Деренко М.В., Денисов Г.А., Малярчук Б.А. и др. // Генетика. 2001. Т.37. №10. С.1402—1410.
9. Степанов В.А., Харьков В.Н., Солтобаева Ж.О. и др. // Генетика. 2001. Т.37. №2. С.256—259.
10. Derbeneva O.A., Starikovskaya E.B., Wallace D.C., Sukernik R.I. // Am. J. Hum. Genet. 2002. V.70. P.1009—1014.
11. Sajantila A., Lahermo P., Anttinen T. // Genome Res. 1995. V.5. P.42—52.
12. Brown M.D., Hosseini S.H., Torroni A. et al. // Am. J. Hum. Genet. 1998. V.63. P.1852—1861.
13. Derenko M.V., Grzybowski T., Malyarchuk B.A. et al. // Am. J. Hum. Genet. 2001. V.69. P.237—241.
14. Рычков Ю.Г., Перевозчиков М.В., Шереметьева В.А. и др. // Вопр. антропологии. 1969. Вып.31. С.3—32.
15. Santos F.R., Pandya A., Tyler-Smith C. et al. // Am. J. Hum. Genet. 1999. V.64. P.619—628.
16. Karafet T.M., Zegura S.L, Posukh O. et al. // Am. J. Hum. Genet. 1999. V.64. P.817—831.
17. Rosser Z.H., Zerjal T., Hurles M. et al. // Am. J. Hum. Genet. 2000. V.67. P.1526—1543.