
№9, 2007 г.
©Алифанов В.Р.
Ящерицы в эпоху динозавров
В.Р. Алифанов
Владимир Рудольфович Алифанов, кандидат биологических
наук, старший научный сотрудник Палеонтологического института РАН.
Реликты юрского периода
На фоне современного разнообразия ящериц, достигающего нескольких тысяч видов, палеонтологическая летопись выглядит бедно. И не удивительно! Останки небольших животных обычно быстро разрушаются и редко сохраняются. Тем не менее благодаря находке нескольких мелких челюстных костей в нижнемеловых отложениях Англии в середине XIX в., т.е. на заре развития палеонтологии позвоночных, выяснилось, что ящерицы были современниками динозавров. Но еще раньше появились сведения о гигантских водных ящерицах - мозазаврах, обитавших в морях на последнем этапе мезозойской эры.
Со времени первой находки на территории Голландии в 1780 г. особенности образа жизни и родства мозазавров не раз обсуждались в среде специалистов. В их числе были такие известные палеонтологи, как Ж.Кювье, Р.Оуэн, Э.Коп, О.Марш и Л.Долло. В итоге удалось установить, что возникновение и расцвет морских ящериц - всего лишь яркий эпизод сложного и длительного исторического развития в целом наземной группы, которая, как сейчас доподлинно известно, появилась на арене эволюции в середине юрского периода, т.е. около 170 млн лет назад.
Начало распространения ящериц обусловлено рядом причин, в том числе тем, что в течение мезозойской эры на планете был относительно ровный и теплый климат, без оледенений и со слабо выраженной широтной зональностью. Это весьма важное обстоятельство не только для ящериц, но и для всех пресмыкающихся, поскольку их активность, в отличие от птиц и млекопитающих, зависит от внешнего тепла. С одной стороны, подобная физиологическая особенность позволяет экономно расходовать энергию, поступающую с пищей, а с другой, ставит в зависимость от суточных или сезонных температур. Чем холоднее или чем больше температурные колебания, тем труднее пресмыкающимся закрепиться на той или иной территории. Вот почему в мезозое они легко расселялись во всех направлениях, а современная герпетофауна сосредоточена главным образом в субтропических и тропических широтах.

Скелет хайнозавра - одного из многих видов мозазавров.
Эти морские ящерицы были самыми страшными хищниками морей в конце мезозоя.
Некоторые из них, как хайнозавр, достигали 15-17 м в длину.
Во многом эволюционный взлет ящериц сопряжен с рядом физиологических и морфологических новоприобретений. Наиболее важные из них касались строения черепа. В результате верхняя челюсть ящериц приобрела подвижность, а челюстной аппарат в целом получил возможность амортизировать рывки добычи, перехватывать ее для максимально выгодного приложения силы челюстей и быстро проталкивать в глотку. Расчеты показывают, что подвижный, или кинетический череп (у ящериц черепная конструкция называется амфикинетической из-за наличия двух поворотных зон в крыше черепа) максимально эффективен, если добыча невелика и не требует какой-либо специальной обработки челюстями. По этим показателям для ящериц наилучшим образом подходят насекомые [1], ставшие исключительно многочисленными в начале мезозоя.

Сцена охоты мозазавра.
Пищей морским ящерицам служила небольшая рыба,
но они вступали в схватки и с разнообразными врагами,
в том числе и с гигантскими кальмарами.
Причуды летописи
Научный интерес к ящерицам проявился на рубеже XVIII-XIX вв. Тогда специалисты, знакомясь с их разнообразием, опирались главным образом на внешние характеристики. Во второй половине 19-го столетия, когда начали распространяться эволюционные идеи, возникла необходимость создать филогенетическую классификацию ящериц, для чего потребовались данные сравнительной анатомии. На этом этапе палеонтология не оказала заметного влияния, поставив в число основных лишь вопрос о происхождении и родственных связях мозазавров.
В конце XIX-начале XX в. появилось сразу несколько развернутых классификаций. Однако в основу современной картины легла система американского ученого Ч.Кэмпа [2]. В ней фигурируют названия принятых и сейчас крупных систематических образований (инфраотрядов): сцинкоморфы (сцинки и их родственники), ангвиморфы (вараны, ядозубы, веретеницы и их родственники), геккоты (гекконы и чешуеноги) и игуании (агамы и игуаны).
Кэмп указывал на близкое родство сцинкоморф и ангвиморф, а также геккот и игуаний, причем вторую пару считал архаичнее первой. В последние десятилетия эти представления не раз подвергались переосмыслению. В результате геккоты были сближены со сцинкоморфами и ангвиморфами в составе общей группы сцинко-геккономорф, а статус наиболее близких к предковому типу ящериц сохранен только за игуаниями [3, 4]. Но последнее заключение не бесспорно. Ему, например, противоречат данные палеонтологической летописи, в которой первыми о себе заявляют сцинко-геккономорфы.
Впрочем, у палеонтологической летописи есть свои причуды. Например, еще недавно бытовало представление о кайнозойском расцвете ящериц на том основании, что преобладающее число их ископаемых находок обнаружено в отложениях соответствующего возраста. Однако в последние десятилетия все отчетливее стал прорисовываться мезозойский подъем разнообразия группы. Этому содействовало изучение многочисленных материалов из континентальных отложений пустыни Гоби, разнообразных систематически и уникальных по сохранности. В Монголии ископаемые ящерицы собирались и обрабатывались участниками нескольких международных палеонтологических экспедиций: Польско-Монгольской (1964-1972 гг.), Советско-Монгольской (с 1969 г.) и Монголо-Американской (с 1992 г.).
Но первые сборы были сделаны в начале 20-х годов XX в. экспедицией Американского музея естественной истории (Нью-Йорк) на верхнемеловом местонахождении Баин-Дзак. В печати их представил американский палеонтолог Ч.Гилмор [5]. Из нескольких фрагментарных образцов наиболее примечательным выглядит макроцефалозавр (Macrocephalosaurus), череп которого оказался на редкость, даже для современных форм, внушительного размера - более 10 см в длину (этим и объясняется его название: большеголовая ящерица). Любопытна еще одна деталь: на передних концах верхних челюстей макроцефалозавра имелись крупные клыкообразные зубы, как у современных агам. Эта и некоторые другие особенности склонили ученого к выводу о связи макроцефалозавра именно с агамообразными ящерицами.
Сборы других экспедиций существенно расширили представление о разнообразии ископаемых ящериц Азии, но одновременно обнажили новые проблемы. В частности, оказалось, что многие меловые формы не поддаются однозначному систематическому определению. Так, польский палеонтолог А.Сулимский предположил, что установленный им по полному черепу адамизавр (Adamisaurus) находится в тесном родстве с агамами [6]. Действительно, такой вывод можно сделать, опираясь на строение, скажем, костей нижней челюсти. Однако спустя несколько лет Сулимский решил, что сходство адамизавра и агам поверхностно, а гипотезу родства ископаемой формы необходимо пересмотреть в пользу сцинкоморф [7]. Польский палеонтолог описал также несколько новых видов макроцефалозавров [8]. Как оказалось, их “заклыковые” зубы имели лопаткообразную вершину, разделенную на многочисленные дополнительные зубчики, как у некоторых современных игуан. Но эти детали исследователь счел незначительными и, как и адамизавра, отнес макроцефалозавров к сцинкообразным ящерицам.

Череп адамизавра (Adamisaurus magnidentatus).
По форме зубов он напоминает некоторых сцинко-геккономорфных ящериц,
но строением нижних челюстей близок к агамидам.
Образец из фондов Палеонтологического института (ПИН).
Проблему родства адамизавра и макроцефалозавров попытался решить американский палеонтолог Р.Эстес [9]. Ему принадлежит идея включить этих ископаемых ящериц в состав тейид (Teiidae) - семейства, представители которого ныне населяют главным образом Южную Америку, а в меловое время были распространены по всей Америке. Интересно, что положение этой группы, состоящей из двух подгрупп (макро- и микротейин), в системе ящериц точно не установлено. Некоторые систематики допускают, что макротейины, которых еще называют американскими варанами, родственны игуаниям. Однако общепринятая, но не имеющая очевидных морфологических доказательств точка зрения указывает на принадлежность этой группы сцинкоморфам.
Противоречия в суждениях специалистов имеют свои причины. Главная из них в том, что действующая классификация, созданная преимущественно на основании сравнительноанатомического изучения современных форм, не всегда отвечает древнему разнообразию. Вместе с тем именно по находкам ископаемых животных удается вскрыть заблуждения, неточности и противоречия многих сложившихся ранее взглядов. И хотя изложение естественной истории ящериц в наше время уже трудно представить без учета палеонтологического материала, но сказать, что он лежит в основе современных представлений об эволюции этой группы пресмыкающихся, еще нельзя.
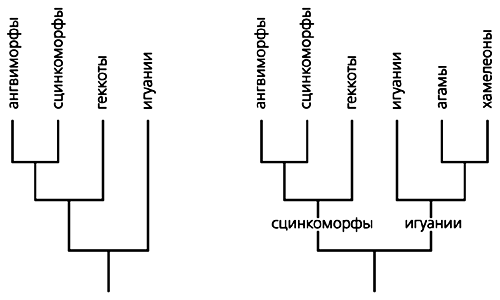
Варианты родственных связей основных подгрупп ящериц. Слева - вариант, предложенный Р.Эстесом и др. [4], справа - автором [14].
Игуаны и историческая зоогеография
Открытие в составе комплексов позднемеловых тетрапод Азии игуан (Iguanidae) современного типа для многих герпетологов стало неожиданностью [10]. Дело в том, что в настоящее время ящерицы этой группы распространены в основном в Северной и Южной Америках. Правда, они обитают еще на Мадагаскаре (три вида) и на островах архипелагов Фиджи и Тонга (один вид), что кажется исключением из общего правила. Само же правило гласит: игуаны живут в Новом Свете, а их ближайшие родственники агамы (Agamidae) - в Старом Свете (кроме Мадагаскара) и Австралии. Иными словами: там, где обитают игуаны, нет агам.
Установлением и объяснением закономерностей расселения животных в прошлом занимается историческая зоогеография. Возможности этой дисциплины еще далеко не исчерпаны, о чем можно судить по разночтению гипотез, объясняющих происхождение современной фауны Австралии с ее обилием эндемичных (т.е. обитающих только на данной территории) видов сумчатых млекопитающих. Если принять, что большинство видов плацентарных млекопитающих появилось на этом окруженном со всех сторон морями южном материке вместе с человеком или немного раньше, то можно сделать некоторые любопытные выводы. Один из них: центр происхождения сумчатых территориально был обособлен от центров происхождения других групп млекопитающих. И следующий вывод: австралийская фауна обособилась прежде, чем началось расселение плацентарных.
По представлениям ряда геологов и биологов, Австралия превратилась в остров в меловое время, более 120 млн лет назад, после чего начала двигаться на север. До этого в триасовом периоде, около 220 млн лет назад, материк еще был частью сверхконтинента Пангеи, а затем входил в состав Гондваны, достоверно включавшей Австралию, Антарктиду и Южную Америку. На севере располагался другой суперконтинент - Лавразия, куда входили Северная Америка, Европа и Палеоазия. Видимо, Гондвана и стала родиной сумчатых. Плацентарным же досталась Лавразия. Она, кстати, просуществовала недолго - на границе юрского и мелового периодов разделилась на несколько ограниченных морями блоков.
В течение мезозоя уровень моря постоянно повышался (эпоха трансгрессии), что во второй половине мелового периода привело к изоляции всех крупных участков суши. Единственно достоверное крупное зоогеографическое событие того времени - обмен фаун Палеоазии и Северной Америки через Берингийский мост. Обычно считается, что мост действовал на протяжении большей части позднего мела, однако есть представление и о его краткосрочном существовании. Последняя точка зрения основана на том, что позднемеловые фауны Азии и Северной Америки включают эндемичные группы, которые нередко формируются в условиях территориальной изоляции. С учетом этого обстоятельства нельзя исключить, что впервые азиоамериканская связь начала действовать на рубеже раннего и позднего мела.
Однако формирование Берингии не объясняет всех особенностей распространения наземных животных в меловое время. Например, одни группы динозавров (тираннозавриды, гребнеголовые гадрозавры, высшие рогатые динозавры) заселяли Палеоазию и Северную Америку, а другие (плоскоголовые гадрозавры, альваресзавры, анкилозавры из семейства нодозавров) - еще и Южную Америку. Отсутствие некоторых типичных для северных континентов групп динозавров в Южной Америке можно объяснить двумя способами: или неполнотой палеонтологической летописи, или допущением еще одного крупного зоогеографического события.
Им могла быть межамериканская фаунистическая связь, которая если имела место, то до начала формирования Берингии. Фактически можно предположить поочередность связей Северной Америки: сначала с Южной Америкой, а потом с Палеоазией. Именно такой ход событий позволяет понять, почему ареал азиатских по происхождению динозавров не расширился далее Северной Америки, тогда как их неазиатские группы освоили не только Южную или Северную Америку, но и Палеоазию.
Но вернемся к игуанам. Разгадать тайну их расселения совсем не просто. Р.Эстес считал, что у этой группы исключительно гондванские корни. Позднее она исчезла в Африке и Австралии, но сохранилась в Южной Америке, где также обнаружены ее позднемеловые формы. По мнению Эстеса, именно с территории этого континента на границе мезозоя и кайнозоя игуаны проникли в Северную Америку - предположительно на растительных плотах.
Однако открытие игуан в позднемеловых отложениях Азии подрывает логику Эстеса. Теперь можно допустить распространение группы не с юга на север, а в противоположном направлении, не в конце позднего мела, а в начале, и не на плотах, а по суше. Не лишним будет заметить, что расселяться с помощью плотов могли бы отдельные виды мелких наземных животных, в том числе ящерицы, но никак не динозавры. Для этих довольно крупных животных такой способ явно не подходит. Так или иначе, но родиной игуан могли стать и Северная Америка, и близкая к ней Палеоевропа. Нельзя сбрасывать со счетов и Африку, по некоторым представлениям, еще в юрское время потерявшую связь с южной группой континентов и установившую фаунистическое сообщение с западной частью Лавразии уже после изоляции Палеоазии [11, 12]. В этом случае родиной игуан могла стать и Афроамерия, объединявшая через Европу Африку (с Мадагаскаром) и Северную Америку, а их проникновение (в сопровождении некоторых групп американских динозавров) на территорию Палеоазии произошло вторично через Берингийский мост. Это так еще и потому, что в Азии раннемеловые игуаны не обнаружены.
Палеонтологические данные открывают возможность по-новому взглянуть и на заселение игуанами о-вов Фиджи и Тонга, куда, как считалось, они попали на растительных плотах из Южной Америки. Теперь допустимо предположить, что игуаны проникли на архипелаги из Юго-Восточной Азии, которая, судя по находкам динозавров, в меловое время была полуостровной частью палеоазиатской суши.
Итак, упомянутое правило выглядит результатом довольно случайного сочетания ряда исторически сложившихся обстоятельств. Впрочем, без конкуренции между агамами и игуанами все же не обошлось.
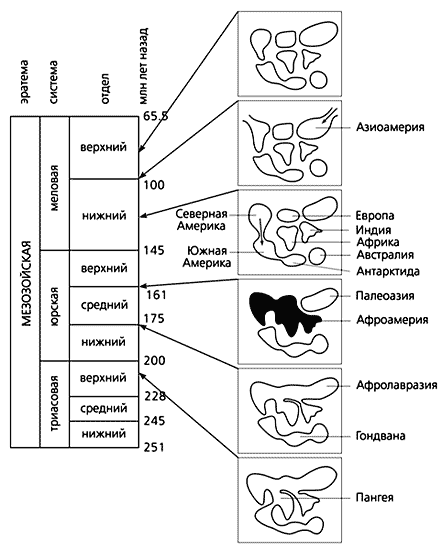
Реконструкция последовательности континентальных связей в мезозое по фаунистическим данным [11, 12].
Афроамерия (выделена черным), возможно, была ареалом первичного распространения игуан
(цветными стрелками указаны главные направления их расселения в меловое время).
Вершина эволюции
В начале позднего мела центральная часть Палеоазии представляла собой обширную низменность, куда ветер и вода сносили большие массы песка. В сезоны дождей здесь появлялось множество временных водоемов, берега которых быстро зарастали. На мелководьях и на берегу многочисленными становились беспозвоночные: черви, рачки, насекомые и их личинки. Ими кормились мелкие млекопитающие, птицы и ящерицы. Однако эти позвоночные и сами легко могли попасть в челюсти хищных динозавров - теропод, среди которых преобладали дромеозавры, имевшие, судя по новейшим данным, птицеообразный облик. Из них больше других известен двухметровый велоцираптор (Velociraptor). У него, как и у всех его ближайших сородичей, была небольшая голова, длинные и узкие челюсти, крупные передние лапы и увеличенный в размере коготь на втором пальце стопы.
Тому, что тероподы не упускали случая схватить и проглотить ящерицу, есть неопровержимое доказательство - плита литографских сланцев, на поверхности которой виден практически полный скелет компсогната (Compsognathus) с остатками баваризавра (Bavarisaurus) в области желудка. Трапеза динозавра, жившего в центре Европы, случилась в конце юрского периода (около 150 млн лет назад).
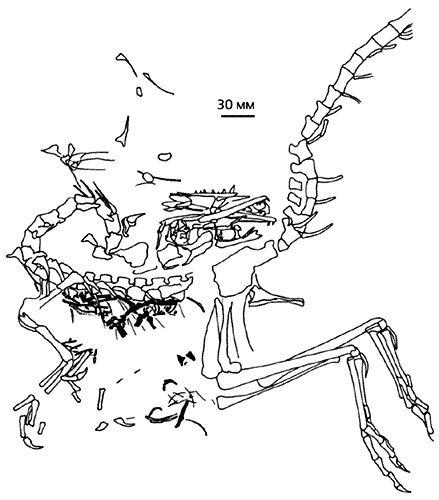
Скелет динозавра компсогната из верхнеюрских сланцев Баварии
с остатками ящерицы (выделены черным цветом) баваризавра в области желудка [16].
Существовали и такие ящерицы, встреч с которыми велоцирапторы избегали. Это крупные двухметровые эстесии (Estesia). С ними без труда могли справиться разве что тираннозавры. Но динозавры-гиганты сторонились гобийских низин, затапливаемых в сезон дождей, оставляя на вершине пищевой пирамиды крупных вараноидных ящериц. Они, конечно, проигрывали в скорости более маневренным и активным тероподам, но мощь и способность плавать позволяла им успешно конкурировать за пространство и пищевые ресурсы.

Силуэты варана эстесии (слева) и динозавра велоцираптора.
Мощь и способность плавать позволяли крупным варанам
успешно конкурировать с динозаврами за пространство и пищевые ресурсы.
Сейчас среди ящериц на положении доминирующих хищников находятся только вараны о.Комодо, длина тела которых доходит до 3 м. Примерно миллион лет назад в Австралии ту же роль играли семиметровые мегалании. Гигантизм нетипичен для ящериц, но может сформироваться при каких-то особых обстоятельствах. Судя по условиям обитания комодского дракона и мегалании, можно предположить, что увеличение размеров стимулируется обилием пищи и отсутствием конкуренции.
Говоря о варанах, нельзя не вспомнить о том, что еще со времен Кэмпа они считаются предками змей. Эта гипотеза, получившая название платинотной (от Platynota - названия вараноидных ящериц в классификации Кэмпа), основана на отдельных признаках сходства двух групп. В числе таких черт упоминаются, например, длинный и подвижный язык; крупные капсулы якобсонова органа, отвечающего за хеморецепцию; острые зубы; дополнительное подвижное сочленение ветвей нижней челюсти; обширные мыщелки (выступы) тел позвонков. На самом деле все эти особенности вместе свойственны лишь представителям современных варанид (Varanidae), тогда как у их ископаемых и современных родственников сходство со змеями практически не выражено. Скорее, все перечисленные признаки произошли параллельно в связи с переходом варанов и змей на питание небольшими позвоночными. Кстати, челюстной аппарат вараноидных сохраняет типичные для ящериц особенности, хотя снабжен сравнительно более мощной мускулатурой. Совсем иначе обстоит дело с челюстным аппаратом змей.

Грудные позвонки современного серого варана (слева), достигающего 1.5 м в длину,
и ископаемой вараноидной ящерицы (Cherminotidae), превышавшей 2 м (верхний мел Монголии).
Экз. ПИН, №4216/206.
Пищевая специализация большинства змей направлена на проглатывание крупной добычи. Это обеспечивается высокой взаимной подвижностью и растяжимостью некоторых внутричерепных соединений, одновременно ведущих к потере силы челюстей. Вот почему многие змеи, чтобы подавить сопротивление жертвы, предварительно ее душат или кусают, впрыскивая яд. Таким образом, специализация челюстного аппарата змей и ящериц различна, и лишь по ней связать две группы отношением предки-потомки затруднительно.
Кстати, змеи, появившиеся в раннемеловое время в Северной Африке, стали обычным элементом комплексов позвоночных только в кайнозое. В мезозойских отложениях Азии пока не найдено ни одной ископаемой змеи. Данная деталь отчетливо иллюстрирует, что меловые ящерицы не испытывали противодействия со стороны их самых опасных конкурентов.
Пора расцвета
Древнейшая азиатская ящерица - чангетизавр (Changetisaurus) из среднеюрских отложений Киргизии. Ее относят к парамацеллодидам (Paramacellodidae), вымершему семейству сцинкоморф. Парамацеллодид находят в позднеюрских, иногда в раннемеловых отложениях всех северных континентов, как и дорсетизаврид, вероятно, принадлежавших ангвиморфам.
Многочисленными ящерицы становятся во второй половине раннего мела вместе с началом периода потепления, состоявшегося примерно 120 млн лет назад. Изменения отразились и на разнообразии других групп позвоночных, в том числе динозавров. О расцвете ящериц того времени лучше всего можно судить по находкам из монгольского местонахождения Хобур, расположенного вблизи юго-восточной оконечности Хангайского хребта. В комплексе представлен десяток семейств, в основном вымершие группы сцинко-геккономорф. Есть в нем парамацеллодиды и дорсетизавриды. Единственная достоверно определяемая современная группа в Хобуре - гекконы [13].

Остатки раннемеловых ящериц из местонахождения Хобур.
В основном это разрозненные элементы скелета,
среди которых преобладают челюстные кости сцинко-геккономорф.
К началу позднего мела некоторые прежние семейства ящериц вымерли, но появилось много новых групп, часть из которых мигрировала из Северной Америки через Берингийский мост. Только на юге Монголии из нескольких верхнемеловых местонахождений известно 18 семейств. Такого не знает ни один регион ни в древности, ни в современности. Пожалуй, этот показатель, как никакой другой, свидетельствует об истинном времени расцвета ящериц.
Основное разнообразие позднемеловых ящериц древней Азии составляли игуании. Здесь были представлены не только все филогенетические ветви этой группы, но и близкий к современному набор жизненных форм. Так, макроцефалозавры, принадлежавшие, судя по строению нижней челюсти, агамо-хамелеоновому стволу игуаний, нередко напоминали современных крупных игуан, но обладали “клыками”. У некоторых из большеголовых ящериц, как, например, у апризавра (Aprisaurus), их было несколько пар. Однако другие макроцефалозавры внешне походили на макротейин. Таков был джадохтозавр (Dzhadochtosaurus). Собственно макротейины тоже обнаружены в комплексах древней Азии, но эти ящерицы были там редки.
Следует сказать и о большом разнообразии так называемых прискагам, которые внешним видом и строением зубной системы напоминали агам с характерными для них “клыками” и уплощенными субтреугольными заклыковыми зубами. Однако ближайшие современные родственники прискагам - хоплоцерк (Hoplocercus) и моруназавр (Morunasaurus) - живут в Южной Америке и считаются игуанами. А вот у настоящего родственника агам, изодонтозавра (Isodontosaurus), зубы лишены явных признаков сходства с зубами современных представителей, в том числе и по характеру прикрепления.
Обычно зубную систему агам и хамелеонов относят к акродонтному типу, при котором зубы занимают на челюстных костях верхушечное положение. Иногда ящериц этих групп даже называют акродонтными игуаниями. Особый статус акродонтности придал американский ученый Э.Коп еще в XIX в. С тех пор термин прочно устоялся в литературе как один из трех основных способов прикрепления зубов у наземных позвоночных наряду с текодонтным (зубы сидят в углублениях - теках, характерен, например, для крокодилов) и плевродонтным (зубы прирастают к челюсти сбоку, как у большинства ящериц). Однако критический взгляд на особенности строения зубной системы акродонтных игуаний показывает, что зубы, которые закладываются на эмбриональных стадиях онтогенеза, прочно срастаются с челюстными костями и перестают расти. По мере увеличения индивидуальных размеров животных эмбриональные зубы становятся миниатюрными, сохраняя свое положение вблизи кромок челюстей, т.е. становятся акродонтными. Однако исходно их тип прикрепления плевродонтный, как у “клыков” (если они выражены) или постэмбрионально возникших зубов. Таким образом, типологическая особенность зубной системы акродонтных игуаний выглядит преувеличенной, что, как выясняется, привело и к недооценке разнообразия этой группы, в том числе ископаемого. Кстати, у изодонтозавра нет “клыков”, эмбриональные и постэмбриональные зубы почти не отличаются друг от друга морфологически, а вот процесс сменности явно ограничен.

Черепа некоторых позднемеловых игуаниевых ящериц Монголии:
1 - апризавра (Aprisaurus bidentatus, экз. ПИН №3142/302);
2 - джадохтозавра (Dzadochtosaurus giganteus, экз. ПИН №3143/103);
3 - хамелеогната (Chamaeleognathus iordanskyi, экз. ПИН № 3142/345);
4 - изодонтозавра (Isodontosaurus gracilis, экз. Ин-та палеобиологии Польской АН, MgR II/39);
5 - параварана (Paravaranus angustifrons, экз. ПИН № 4487/15).
Новые палеонтологические находки в меловых отложениях Монголии показывают, что проблема разнообразия древних и ископаемых игуаний становится исключительно актуальной. И тут возможны некоторые неожиданные решения и подходы. Например, рассмотрения требует основательно забытая и выпавшая из обсуждения гипотеза о родстве с игуаниями мозазавров. Пока преобладает представление о связи мозазаврообразных ящериц с вараноидными ящерицами и со змеями, которая аппелирует к некоторым общим и возникшим вторично признакам черепа.
Кстати, к мозазаврообразным ящерицам из меловых отложений Монголии относится параваран (Paravaranus). Как и у типичных морских ящериц, у него отмечаются удлиненные носовые отверстия и узкая лобная кость. Сближают параварана с мозазавридами и некоторые общие черты в строении костей крыши черепа и нёба. Однако в отличие от морских ящериц параваран отличался миниатюрными размерами и, скорее всего, вел наземный образ жизни.
Итак, разнообразие мезозойских ящериц выглядит довольно внушительно.
Важнейшим итогом его изучения стало представление об исключительном видовом
богатстве игуаний в прошлом и о том, что таксономический ранг самой группы
не уступал таковому сцинко-геккономорф [14].
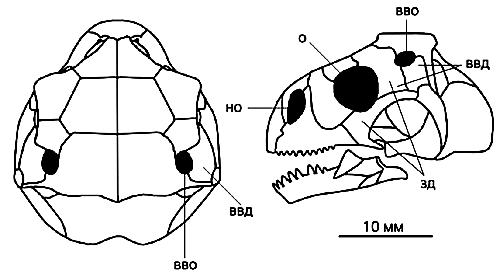
Реконструкция черепа (вид сверху и сбоку) синеоамфисбены (Sineoamphisbaena hexatabularis).
В отличие от других амфисбен у нее есть заглазничные (зд) и верхние височные дуги (ввд).
Черным цветом показаны носовые отверстия (но), орбиты (о) и верхние височные окна (вво).
Говоря о меловых ящерицах древней Азии, нельзя обойти вниманием синеоамфисбену
(Sineoamphisbaena), описанную в литературе по находкам в верхнемеловых
отложениях Внутренней Монголии (Китай) [15]. Широкая
костная межглазничная перегородка, крупная мозговая капсула, раздвоенный
затылочный мыщелок и укороченная нижняя челюсть и ряд других признаков
сближают эту ископаемую форму с амфисбенами, или двуходками. Современные
амфисбены ведут скрытный образ жизни - обитают в норах, которые роют головой.
Видимо, с этим связаны такие преобразования, как утрата конечностей, редукция
некоторых костей крыши черепа. Однако в отличие от раннекайнозойских и
современных амфисбен, у синеоамфисбены были хорошо развиты передние конечности
(задние не обнаружены), а элементы верхних височных дуг напоминали эти
же детали строения у ящериц, хотя и имели некоторые ограничивающие амфикинетизм
особенности. И то, и другое свидетельствует об архаичности ископаемой формы,
еще не достигшей совершенства в роющей специализации. Похоже, что в образе
синеоамфисбены проявляются черты несомненного родства амфисбен и ящериц.
Но включать первых в состав вторых нет необходимости.

Череп и передняя часть скелета славойи (Slavoia darevskyi), архаичной сцинкоморфной ящерицы из верхнемеловых отложений Монголии.
У славойи парадоксально сочетаются миниатюрные глазницы и хорошо развитые конечности. Экз. ПИН № 4487/14.
Следует заметить, что в разных филогенетических стволах сцинко-геккономорф существуют формы, которые воспроизводят те или иные морфологические особенности амфисбен. Нередко считается, что подобного рода сходства возникли вторично, а первичной была наземная специализация. Однако не исключено, что исходно ящерицы вели скрытный образ жизни. В дальнейшем одни их группы освоили подземные биотопы, утратив конечности и приобретя способность рыть норы головой, а другие приспособились к наземной жизни, усовершенствовав способы передвижения и развив зрение и слух. Примером “древнего” типа может служить сцинкоморфная славойя (Slavoia), которая в позднем мелу Монголии была наиболее массовой. У этой миниатюрной ящерицы сочетается, как кажется, несовместимое - незначительные размеры глазниц (т.е. крошечные глаза), позволительные подземным жителям, и хорошо развитые конечности, столь необходимые для наземного передвижения.
В эпоху перемен
По крайней мере за несколько миллионов лет до конца мезозоя разнообразие азиатских ящериц сократилось больше чем в четыре раза. Тогда число семейств снизилось с 18 до четырех. Был еще один и более поздний кризис, который выявляется по составу раннекайнозойских групп: меловые корни только у одной из них. Таким образом, неуклонный рост разнообразия в течение юры и мела неожиданно сменился быстрым падением.
Ключом к пониманию главной причины кризиса может послужить раннекайнозойская история группы.
В начале кайнозоя в центральной части Азии ящерицы вновь оказались «на подъеме», но за счет отсутствующих в прежнее время семейств. Видимо, их заселение происходило вторично, с соседних территорий. Судя по анализу систематического состава раннекайнозойских комплексов, процесс протекал не однократно, а волнообразно в течение палеогенового периода. Первая волна пришла с северо-востока, через Берингию, из Северной Америки и связанной с ней в начале кайнозоя Европы. Вторая - с юга, из Индии, присоединившейся к Азии после меловой изоляции. Третья - западная, или европейская, - возникла после того, как Европа утратила связь с Северной Америкой, а отделявшее Европу от Палеоазии во второй половине мезозоя и в начале кайнозоя Тургайское море высохло. Благодаря северо-восточной волне появились агамы, близкие к современной агаме-бабочке (Leiolepis). С южной волной прибыли неядовитые змеи, вараны современного типа и агамы, схожие с современным шипохвостом (Uromastyx). Западная волна принесла в Центральную Азию ящериц тех родов и семейств (в том числе настоящих ящериц - Lacertidae), которые сейчас обычны в умеренных широтах Евразии.
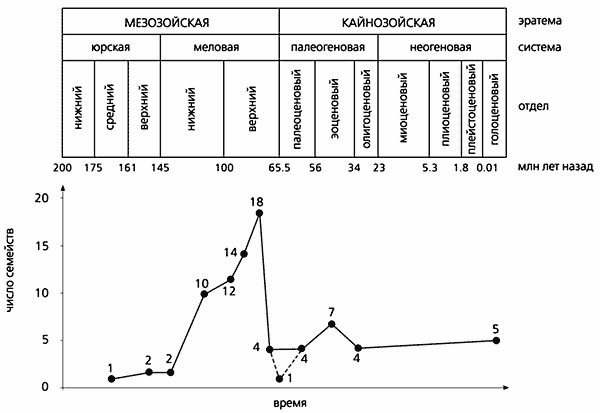
Динамика разнообразия семейств ископаемых ящериц в
мезозое и в кайнозое.
В кайнозое наибольшее и наименьшее число семейств
совпадают с периодами самых высоких и самых низких палеотемператур.
Вероятно, этим же объясняются и мезозойские максимумы
и минимумы разнообразия.
Примечательно, что зависимые от условий окружающей среды ящерицы в кайнозое, как и в мелу, распространялись через высокоширотный Берингийский мост. Это связано с очередным потеплением, приведшим в середине первой половины кайнозойской эры (в эоцене) к установлению самого высокого для того времени температурного максимума. На фоне потепления число семейств ящериц увеличилось с четырех до семи. Однако к концу эоцена разнообразие центральноазиатских ящериц вновь упало. Причина - хорошо известное похолодание. Оно же прямо или опосредованно привело даже к обеднению комплексов доминирующих в кайнозое млекопитающих. Скорее всего, позднемеловой кризис азиатских ящериц и рептилий в целом - тоже следствие одного или серии похолоданий.
Как бы то ни было, но, в отличие от динозавров, ящерицы дожили до наших дней. Спасительными для них стали небольшие размеры, способные обеспечить быструю смену поколений, облегчить приспособляемость к разным биотопам и компенсировать потери численности и разнообразия в послекризисные времена. Вероятно, именно этих качеств не хватило динозаврам в эпоху глобальных перемен на рубеже мезозойской и кайнозойских эр.
Работа выполнена в рамках программы
Президиума РАН 25 “Происхождение и эволюция биосферы” на 2005 г. (подпрограмма
2)
и поддержана грантом Президента
РФ для Ведущей научной школы НШ-6228.2006.4.
Литература
1. Иорданский Н.Н. Эволюция комплексных адаптаций. Челюстной аппарат амфибий и рептилий. М., 1990.
2. Сamp C.L. // Bull. Am. Mus. Nat. Hist. 1923. V.48. P.289-481.
3. Суханов В.Б. // Зоол. журн. 1961. Т.40. №1. С.73-83.
4. Estes R., de Queiroz K., Gautier J. Phylogenetic relationships within Squamata // Phylogenetic relationships within lizards families / Eds R.Estes, G.K.Pregill. Stanford, 1988. P.119-281.
5. Gilmore C.W. // Bull. Amer. Museum Natur. Hist. 1943. V.81. P.361-384.
6. Sulimski A. // Palaeontol. Polonica. 1972. V.27. P.33-40.
7. Sulimski A. // Palaeontol. Polonica. 1978. V.38. P.43-56.
8. Sulimski A. // Palaeontol. Polonica. 1975. V.33. P.25-102.
9. Estes R. Sauria terrestria, Amphisbaenia // Handbuch der Palaeoherpetologie. Stuttgart, 1983. T.10A.
10. Borsuk-Bialynicka M., Alifanov V.R. // Acta Palaeontol. Polonica. 1991. V.36. №3. P.325-342.
11. Каландадзе И.И., Раутиан А.С. Система млекопитающих и историческая зоогеография // Филогенетика млекопитающих. М., 1992. С.44-152.
12. Kalandadze N.N., Rautian A.S. Historical zoogeography of terrestrial tetrapods and new method of global palaeogeographical reconstructions // Evolution of the Biosphere / Eds Rozanov A.Yu., Vickers-Rich P., Tassel C. Launceston, 1997. P.95-98.
13. Алифанов В.Р. // Палеонтол. журн. 1999. №1. С.124-126.
14. Алифанов В.Р. Макроцефалозавры и ранние этапы эволюции ящериц Центральной Азии. М., 2000.
15. Wu X. -Ch., Brinkman D.B., Russell A.P. // Can. J. Earth Sci.1996. V.33. №4. P.541-578.
16. Evans S.E. // N. Jb. Geol. Palдont. Abh. 1994. Bd.192. №1. P.37-52.
