№9, 2006 г.
© Шейман И.М., Сахарова Н.В.
История из жизни
замечательных червейИ.M. Шейман, Н.В. Сахарова
Инна Моисеевна Шейман, к.б.н., с.н.с. Ин-та биофизики клетки РАН.
Наталия врьевна Сахарова, к.б.н., в.н.с. Ин-та теор. и экспер.биофизики РАН.Замечательные черви - это планарии. Они впервые обратили на себя внимание натуралистов более двух с половиной веков назад: если маленького червячка разрезали на несколько частей, то из каждой части в скором времени образовывалась новая полноценная особь. На протяжении прошедших с тех пор лет это свойство планарий возбуждало интерес у исследователей разных направлений биологии. Способность регенерировать на самом деле присуща в какой-то степени многим видам животных и разным органам. Но у планарий она выражена в такой демонстративной форме, что делает их привлекательными для экспериментального исследования многих вопросов, связанных с биологией развития. В частности, на протяжении длительного времени, включая современный период, эти черви служат излюбленным объектом для изучения основ регенерации. Увлеченные писатели от науки, переносившие описание замечательных особенностей планарий из книги в книгу, утверждали, что червяка можно разрезать на 100 частей, и из каждой части восстановится новое существо. На каком-то этапе описаний число 100 удвоилось. А затем и утроилось. бти числа не имеют под собой достаточно оснований, хотя регенерационная способность, свойственная планариям, действительно огромна.
Сенсация Мак-Коннелла
По зоологической классификации планарии относятся к типу плоских червей, которые представляют ключевое звено в эволюции по ряду важнейших признаков. У этих червей впервые возникает билатеральная симметрия, впервые происходит концентрация нервных элементов в центральную нервную систему, впервые на головном конце тела формируется головной ганглий («мозг»). К этому следует добавить еще ряд замечательных свойств: способность к бесполому размножению наряду с половым; способность к каннибализму; способность в процессе пищеварения превращать трубку кишечника в единый симпласт (многоядерный слой без межклеточных перегородок) с последующим восстановлением исходной структуры; выполнение кишечником одновременно пищеварительной и распределительной функций; и, наконец, колоссальная морфогенетическая пластичность, лежащая в основе регенерации при восстановлении структуры кишечника в ходе пищеварения, во время бесполого размножения, а также при искусственном повреждении тела.
Две планарии, послужившие для изучения памятного следа:
многоглазая Ijimia tenuis (вверху) и двуглазая Dugesia tigrina.Планарии половых форм - гермафродиты, их органы размножения располагаются в передней части тела. Глотка - мышечный орган, с помощью которого поглощается пища, - расположена на середине брюшной поверхности тела. У разных видов планарий (кстати, их обычные размеры - от нескольких миллиметров до нескольких сантиметров) бывают два простейших глаза на переднем конце тела или множество по его краям.
Научный интерес к планариям пережил несколько пиков. Последний случился в середине прошлого века, точнее, в середине 50-х годов. Американский зоопсихолог Дж.Мак-Коннелл изучал поведение этих червей - вырабатывал у них условные рефлексы на сочетания света и раздражения электрическим током и наблюдал сокращения тела. Он также обучал их выбору направления в Т-образном лабиринте. Мак-Коннелл знал о способности планарий к регенерации, а его личная склонность к фантазии возбудила в нем вопрос: что будет, если разрезать поперек обученную планарию и позволить регенерировать двум половинкам? Задумано - сделано. Обе планарии, восстановленные из одной обученной и разрезанной пополам, через две недели после операции воспроизводили результат обучения, т.е. сохраняли образовавшийся до перерезки памятный след! Поскольку у новых особей память была равноценной, а ганглий оставался только у одной исходной половинки планарии, Мак-Коннелл заключил, что «мозг» тут ни при чем. И приписал сохранение памятного следа недифференцированным клеткам - необластам, которые служат материалом для восстановления утраченных половинок [1].
В ту пору мысли биологической научной общественности были прикованы к молекуле ДНК. Мак-Коннелл решил, что подобно тому, как ДНК кодирует генетическую память, информация о памяти мозга может быть записана на подобных ДНК, но более пластичных молекулах - РНК, которыми богаты необласты. Чтобы это проверить, зоопсихолог прибег к оригинальному способу, используя склонность планарий к каннибализму. Обученных червей накалывал наподобие бифштекса и предлагал их в качестве кандидатов на съедение голодным необученным планариям. Поевшие своих собратьев планарии-реципиенты воспроизвели условные рефлексы, или памятный след, своих жертв (доноров). Как и в предыдущем опыте, произошла передача памятного следа новым структурам. Ученый был окрылен «подтверждением» своей идеи о существовании и переносе «молекул памяти», которые, как он думал, передаются и при регенерации, и при кормлении [2].
Последовали эксперименты с выделением общей РНК из обученных планарий и кормлением ею голодных червей, а также введением ее путем инъекций. Результаты иногда давали положительный эффект.
бти сенсационные опыты и мысли Мак-Коннелла вызвали в широких научных кругах ажиотаж и вскружили многие головы, имевшие весьма малое отношение к предмету. Разные лаборатории принялись обзаводиться планариями и кое-как «обучать» их, резать, кормить разными РНК и т.д.Иногда опыты Мак-Коннелла удавалось воспроизвести, иногда его результаты опровергались. Часто эксперименты ставились неумело. В мировой научной и околонаучной печати шумели дискуссии. Наконец, ажиотаж улегся. Мысли Мак-Коннелла не были ни доказаны, ни опровергнуты, и, главное, результаты многолетних опытов не получили объяснения. Американский зоопсихолог умер под натиском критики, а его сотрудники разбрелись по другим лабораториям.
На родной почве
Отечественные биологи не остались в стороне от описанного ажиотажа. На начало 60-х годов пришлась организация Научного центра биологических исследований в Пущине-на-Оке. В этом Центре предполагалось развивать новые идеи на основе современных подходов и методов преимущественно физико-химической направленности. Заместителем директора-организатора был назначен работавший до того сотрудником Отдела науки ЦК А.Н.Черкашин (в ранней молодости Ч физиолог, ученик академика Л.А.Орбели). В Научном центре планировалось создать Институт проблем памяти. бта идея, в частности, была навеяна «планарной» сенсацией и достижениями молекулярной биологии. Дело в том, что работы Мак-Коннелла также стали причиной возросшего во всем научном мире интереса к явлениям памяти. Он коснулся и организаторов Центра. Имея известную свободу действий и будучи человеком увлекающимся, Черкашин решил тоже принять участие в исследовании памяти на планариях. С этой целью он взял на работу несколько молодых биологов: Инну Шейман, только что защитившую кандидатскую диссертацию по высшей нервной деятельности; бмму Сергееву, окончившую -альневосточный университет, находившийся во Владивостоке; Галю Богоровскую - студентку кафедры эмбриологии МГУ. В составе организованной в Москве первой научной группы биологи прибыли вместе с планариями в Пущино 28 марта 1964 г. Спустя годы группа постепенно пополнилась, в нее пришли: гистолог Иван Ефимов, эмбриологи Наташа Сахарова, Алик Азарашвили и Гарик Тирас, зоологи Люда Попкова и Наташа Крещенко, специалистка по высшей нервной деятельности Елена Зубина.
Из пущинских научных учреждений тогда строился Институт биофизики. Группа разместилась в здании его будущего вивария и приступила к своим первым экспериментам в весьма «романтических» условиях: при отсутствии оборудования, инфраструктур, удобств, магазинов и отрезанности от ученого мира весенним половодьем Оки.
Первые планарии были привезены с биологической станции МГУ, которая находилась в Чашникове. Предстояло найти местные источники животных. Походы по берегам Оки и обследования многочисленных мелких и средних старичных озер оказались успешными - молодые ученые обнаружили планарий нескольких видов. (В более поздние годы сельское хозяйство полностью истребило эту живность, да и уничтожило сами озера.) И приступили к исследованиям.
Для начала повторили опыты Мак-Коннелла. Молодой дух противоречия, сдобренный советским патриотизмом, настроил психологическую «установку» на опровержение результатов, полученных американцем. Каково же было удивление честных энтузиастов, когда эти результаты подтвердились!
И тогда начались поиски научного объяснения сенсационных эффектов. -ля экспериментального анализа придумали два специальных подхода и применили дополнительные методы.Первый подход был использован, чтобы понять, как сохраняется памятный след при регенерации. Планарий перерезали таким образом, чтобы в одних частях тела оставались участки центральной нервной системы (ЦНС), ганглия или продольных нервных стволов, а в других ничего этого не было. Если планарий предварительно обучали, то после регенерации фрагментов с сохраненными элементами ЦНС у новых особей воспроизводился памятный след, а фрагменты, полностью лишенные участков ЦНС, не только его не сохраняли, но и не регенерировали, хотя оставались какое-то время живыми. Так была установлена зависимость регенерации и сохранности памятного следа от наличия ЦНС, хотя бы небольшой ее части. Кроме того, обнаружилось, что после перерезки обученных планарий пополам головная половинка, содержащая ганглий, воспроизводила памятный след даже на следующий день после операции. У хвостового же фрагмента, лишенного ганглия, но с уцелевшей частью ЦНС в виде остаточных нервных стволов, памятный след проявлялся только после пяти суток регенерации.
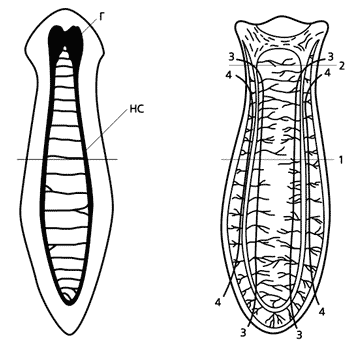
Схема строения центральной нервной системы планарий (слева) и перерезки червя пополам.
Г - ганглий; НС - нервные стволы.Следовало изучить, что происходит в хвостовой половинке планарии за то время, в течение которого памятный след кажется утраченным, но на самом деле сохраняется и обнаруживается позже. Галя Богоровская в своей дипломной работе провела гистологический анализ регенерирующих хвостовых фрагментов в каждый из пяти дней. В результате выяснилось, что после операции происходят интересные события. В направлении обрезанных остаточных нервных стволов и затем вдоль них к раневой поверхности мигрируют необласты - недифференцированные резервные клетки. Их скопление формирует на раневой поверхности регенерационную почку, или бластему. Одновременно в том же направлении прорастают нервные волокна из клеток остаточных нервных стволов, сохранившихся в хвостовом фрагменте. В бластеме необласты собираются вокруг проросших волокон, которые к пятому дню смыкаются. Таким образом формируется зачаток нового ганглия. Позднее Наташа Сахарова вместе с Ритой Гордон и Гариком Тирасом провели гистохимические исследования (изучили нейросекрецию, холинэстеразную активность) и выяснили, что ко времени смыкания в ганглии проявляются свойства нервного образования. Следовательно, необласты уже дифференцировались и приобрели структурные и функциональные черты нейронов за счет контакта с перерезанными нервными стволами. Приобретая специализированные свойства, ганглий оказался способным воспроизводить также и памятный след, образованный у исходной планарии.
Здесь нужно сказать, что собой представляет воспроизведение памятного следа. Не следует думать, что такое явление можно наблюдать у более высоко организованного существа. Дело в том, что планарии - животные с очень ограниченным потенциалом реакций. На любое раздражение черви реагируют (отвечают) сокращением или поворотом тела. Поэтому при выработке у них условных рефлексов применили особый подход: использовали подпороговые величины условных раздражителей, которые в сочетании с электрическим раздражением вызывали реакции (ответ), т.е. становились сигнальными. По существу повышалась чувствительность нервной системы планарий к условным раздражителям, что и было принято за условный рефлекс. При регенерации такое повышение передавалось новым структурам от остатков нервных стволов наряду с сообщением им основных признаков нейронов.
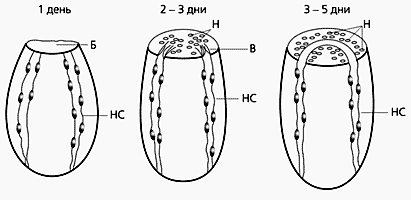
Схема регенерации ганглия планарий.
НС - нервный ствол; В - нервные волокна; Б - бластема,
Н - необласты, дифференцирующиеся в клетки ганглия.Таким образом, передача необластам определенных свойств от сохранившихся нервных стволов была четко установлена. И это объясняет феномен переноса памятного следа новому ганглию при регенерации планарий. (Не исключено, что аналогичный ход событий присущ и более сложным нейрофизиологическим процессам.) Однако механизмы такой «эстафеты», как и направленной дифференцировки необластов, остаются непонятными по сей день.
В течение того времени, пока продолжались целенаправленные исследования, изменялся состав сотрудников группы. В разное время кто-то прибывал, кто-то выбывалЕ Отдалился от нее и главный меценат, что осложнило работу группы.
«Этого не может быть, потому что не может быть никогда»
Второй подход был применен для объяснения наиболее сенсационного «переноса памяти» при каннибализме. Передача памятного следа от старых нервных структур вновь образующимся у регенерирующих планарий имела объяснение на клеточном уровне. Но перенос через каннибализм посредством гипотетических «молекул памяти» действительно представлялся невероятным. Приходила в голову мысль, что сам процесс питания у планарий приводит к общему неспецифическому повышению возбудимости в центральной нервной системе в ответ на любые воздействия. На таком фоне и увеличивается число реакций на предъявляемые раздражители. И эта мысль была верной, поскольку у планарий, как уже говорилось, невозможно получить дифференцированные реакции - на разные раздражители они реагируют качественно одинаково. Выходило, что результаты опытов Мак-Коннелла, а также повторенных у нас, представляют собой артефакты. Чтобы в этом удостовериться окончательно, пришлось изобрести еще один прием.
Начали снова повторять эксперименты Мак-Коннелла. Но наши планарии (Ijimia tenuis), выловленные в старичных озерах вдоль Оки, не желали есть друг друга. Выход нашелся, когда кто-то завез из Америки в Ленинградский университет лабораторную бесполую расу другого вида - Dugesia tigrina. бти черви размножаются простым отделением хвостового фрагмента с последующей регенерацией двух новых особей. Только что пришедший из армии прирожденный биолог Алеша Карпов из нескольких подаренных нам D.tigrina создал огромную популяцию из сотен тысяч планарий. Эти животные охотно поедали своих слегка поврежденных собратьев.
Условные рефлексы вырабатывали у планарий (разделив их на две группы) «хитрым» способом. Каждый из подпороговых раздражителей - вибрацию или освещение - сочетали с электрическим воздействием. Ответ в группах на оба условных раздражителя был одинаковым по форме, но «световые» планарии реагировали только на свет, а «вибрационные» - только на вибрацию. Так образовались две группы обученных планарий. Они и стали жертвами каннибализма - ими покормили планарий-реципиентов. Каждую такую группу в свою очередь поделили пополам и подвергли испытанию на один из условных раздражителей. Итак, половину группы планарий, поевших «световых» доноров, тестировали на свет, а другую половину - на вибрацию. Точно так же поступили с реципиентами «вибрационных» доноров. Вопреки ожиданиям, эффект у реципиентов оказался специфическим, т.е. они отвечали большим количеством реакций только на раздражитель, сигнальный для поеденных ими доноров. Казалось бы, этот опыт подтвердил вывод Мак-Коннелла (во всяком случае, если бы он получил такие данные, то не подвергся бы столь жестокой критике).
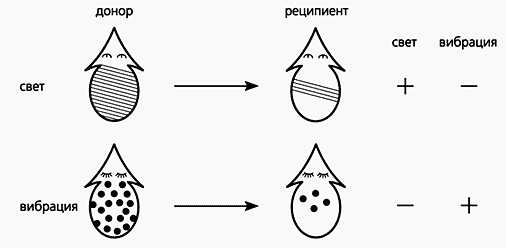
Схема, иллюстрирующая передачу планариям-реципиентам условных рефлексов,
выработанных на свет или вибрацию у планарий-доноров.
Знаками + и - отмечено наличие или отсутствие условного рефлекса у реципиента.Но на этом все не закончилось. Добытые факты были проанализированы во времени. Разные группы реципиентов тестировали условным раздражителем их доноров через разные промежутки времени (часы и дни) после кормления. И тут-то получили новые и совершенно неожиданные результаты. бффект проявлялся через пять-шесть часов после кормления планарий и держался три дня, после чего плавно убывал на четвертые-пятые сутки после кормления. Следовательно, от доноров к реципиентам не переносились «молекулы памяти». Проявлялось хотя и специфическое, но кратковременное повышение возбудимости.
Возникла новая задача. Следовало узнать, как связана полученная картина с процессом, происходящим в теле планарии после кормления, - с пищеварением. Все, что было известно о нем, ни в какой мере не отвечало на вопросы, вызванные последними опытами. Потребовался гистологический анализ тела планарий после кормления по часам и дням, подсказанным предыдущим опытом. За анализ взялась Наташа Сахарова.
Кишечник планарий являет собой трубку с однослойной стенкой, в клетках которой происходит пищеварение. Через пять-шесть часов после кормления эти клетки образуют амебоидные отростки, направленные в сторону внутренних структур червя, в том числе в сторону нервных стволов и ганглия. Известно, что планарии лишены специальной распределительной системы. (У более высоко организованных животных эту функцию выполняет кровеносная система.) Очевидно, что через такие отростки питательные вещества передаются тканям тела после частичного переваривания пищи. В дальнейшем кишечная структура полностью перестраивается, образуется симпласт, в котором «плавают» продолжающие перевариваться пищевые частицы, и который окружает все органы планарий. Там между ними происходит непосредственный контакт. Через три дня начинает восстанавливаться исходная структура кишечника и происходит его естественная регенерация. Пищеварение и доставка клеткам организма питательных веществ протекают одновременно и на этом заканчиваются.
Так была установлена временнАя связь между проявлением следов памяти при каннибализме и процессом пищеварения. Специфические реакции на раздражители воспроизводились у планарий-реципиентов в тот отрезок времени, когда клетки тела (в том числе нервные структуры) контактировали с пищевыми частицами. бто был кратковременный эффект - он исчезал с прекращением пищеварения и, очевидно, с полным распадом пищевых веществ.
Требовалось объяснить довольно сложное - хотя и кратковременное, но специфическое - повышение возбудимости у планарий-реципиентов. И было высказано такое предположение: у планарий-доноров при выработке условных рефлексов повышается возбудимость в тех нервных структурах, которые воспринимают условный раздражитель. При попадании в тело реципиента и частичном распаде осколки этих структур воздействуют на основе биохимического сродства на соответствующие им нервные элементы реципиентов, повышая в свою очередь их возбудимость. После полного распада осколков этот эффект прекращается.
Такое объяснение подтвердилось в сопутствующем опыте, когда применили изотопную метку, чтобы различить двух разных доноров. 3Н-уридин вводили планариям обоих видов (D.tigrina и I.tenuis), реципиентами были планарии D.tigrina. Доноры оказались меченными по-разному: клетки первого вида содержали больше метки, чем второго, а клетки нервных структур меньше, чем других тканей. После кормления такими донорами исследовали распределение метки в теле реципиентов в уже обозначенные в предыдущих опытах временны?е промежутки. В ганглии реципиентов, поевших доноров D.tigrina, метка появилась через 6 час после кормления, а наибольшее число клеток, содержащих 3Н-уридин, было отмечено через трое суток. Затем количество меченых клеток убывало, пока не исчезло полностью. В ганглии планарий, кормленных донорами I.tenuis, меченые частицы распространялись медленнее и менее интенсивно. бти опыты показали, что пищевые частицы разносятся в теле реципиентов активнее, если они одного вида с донорами. Кроме того, подтвердился и неслучайный характер распределения пищевых частиц. Очевидно, они движутся не хаотически, а направленно, и их взаимодействие с клетками реципиентов основано на химическом сродстве. Тем самым подтвердилось высказанное объяснение опытов, в которых поведение обученных планарий передавалось реципиентам.
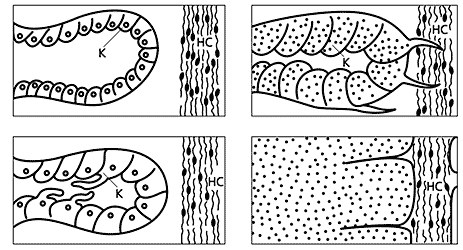
Схема перестройки кишечника.
а - в покое; б - в начале пищеварения; в - через 6 ч после кормления. г - симпласт.
К - клетки стенки кишечника; НС - нервный ствол.Таким образом, обнаружилась корреляция между проявлением специфической возбудимости, клеточной реорганизацией пищеварительной системы и системой активного распределения пищевых веществ. Попортившие много крови нашумевшие опыты по «переносу памяти», наконец, можно было объяснить следующим образом. Специфические вещества, которые образовались при определенной стимуляции доноров и попали в клетки ганглия реципиентов с пищевыми частицами, оказывали возбуждающее действие на соответствующие по химическому сродству их клетки. И у реципиентов имитировалось возбуждение доноров. Это состояние индукции длилось до тех пор, пока не заканчивалось полное переваривание - распад пищевых частиц - и восстановление структуры кишечника.
В печати довольно долго продолжалась критика работ Мак-Коннелла о «молекулах памяти» с известной мотивацией - «не может быть». В газетах выступали философы. Что же касается проведенных нами опытов и высказанных объяснений, то на том же основании нам отказали в публикации. Лишь гистологическую часть удалось малозаметным путем отослать в иностранный журнал [3, 4]. А полностью работа была издана только в канун перестройки [5].
Возвращаясь к морфогенезу
Естественное продолжение этих исследований было заблокировано. Между тем одновременно и в последующих наших экспериментах (в течение 70-80-х годов) на планариях все четче улавливалась связь между процессами, обеспечивающими формирование памятного следа, и элементами морфогенеза. Впервые она обнаружилась, как уже здесь упомянуто, когда анализировался сам факт сохранения памятного следа в процессе регенерации планарии.
Исходя из первоначального интереса к молекулам РНК как переносчикам памяти, у нас была проведена следующая работа. Иван Ефимов помещал планарий в раствор РНКазы, затем перерезал их и на препаратах ганглия наблюдал за естественной деструкцией (апоптозом), сопровождающей любой процесс развития. Спустя четыре часа после перерезки количество разрушающихся клеток в ганглии контрольных и опытных планарий увеличивалось до максимума, который у последних был значительно выше. Если у таких планарий экспериментаторы предварительно выработали условный рефлекс, то он после регенерации не сохранялся. В сходном опыте планарий опускали в раствор РНКазы после перерезки. В этом случае деструкция клеток была ниже, чем у контрольных особей, и сохранялся условный рефлекс. бти опыты никак не объяснили роли РНК в изучаемых процессах, но показали, что такой элемент развития, как деструкция части клеток, существен для сохранения памятного следа [6].
Новое обнаружилось в поведении «факторов регенерации», т.е. агентов, образующихся при восстановительных процессах. Гарик Тирас исследовал, как проявляют себя эти факторы в ходе регенерации планарий. Оказалось, что в разные дни восстановления такие агенты действуют прямо противоположно. Так, экстракты из планарий, регенерировавших в течение одних, двух, трех и пяти суток, при введении другим планариям стимулировали вызванную у них регенерацию. Но если процесс длился четыре дня, экстракт, напротив, тормозил ее. бкстракты из планарий, содержавшие «факторы регенерации», которые стимулировали или тормозили регенерацию, в соответствующих опытах стимулировали или тормозили образование и сохранение условных рефлексов у неповрежденных планарий [7].
Когда сопоставили действие «факторов регенерации» планарий и чужеродного морфогена гидры, который был уже известен и структура которого была идентифицирована как полипептид [8], обнаружилось много общего. Результаты этих опытов побудили нас исследовать влияние некоторых регуляторных нейропептидов позвоночных (адренокортикотропного гормона, люлиберина, их фрагментов) на регенерацию планарий и образование у них памятного следа. В итоге была выделена группа морфогенетически активных нейропептидов, контролирующих регенерацию червей, а также их обучение и сохранение у них экспериментально выработанных рефлексов [9].
Вместо эпилога
Планарии остаются идеальным организмом для исследования клеточных механизмов морфогенеза именно за счет регенерационных способностей. Особое внимание исследователей привлекают к себе необласты - предшественники стволовых клеток позвоночных. В разных лабораториях мира в настоящее время возобновился интерес к планариям как к прекрасному модельному объекту. На них проводятся генетический анализ и микроструктурные работы с применением новейших методов, позволяющих, в частности, прослеживать судьбу маркированных необластов [10].
К сожалению, упомянутые современные работы направлены только на изучение клеточных и молекулярных основ морфогенеза. С планариями больше никто не связывает исследований механизмов памяти. Между тем существуют новые данные о поведении дифференцирующихся клеток и об активации разных генов в ходе регенерации нервных элементов. Использовать такие сведения в работе на простейших моделях обучения или одновременно изучать два важнейших явления (память и морфогенез), применяя новейшие подходы и методы, - какая блестящая и богатая научная перспектива!
Наши же исследования в настоящее время по объективным причинам прерваны. Но это уже совсем другая историяЕ
Литература
1. McConnell J.V., Jacobson A.L., Kimble D.P. // J. Compar. and Physiol. Psychol. 1959. V.52. No.1. P.1-5.
2. McConnell J.V. // J. Neurophysiol. 1962. V.3. Suppl.1. P.542-548.
3. Sheimann I.M., Sakharova N.Yu. // Compar. Biochem. and Physiol. A. 1974. V.48. P.601-607.
4. Sakharova N.Yu., Sheimann I.M. // Compar. Biochem and Physiol. A. 1977. V.58. P.427-431.
5. Шейман И.М. Регуляторы морфогенеза и их адаптивная роль. М., 1984.
6. Шейман И.М., Ефимов И.А., Богоровская Г.И. // Онтогенез. 1971. Т.2. No.4. С.411-418.
7. Тирас Х.П., Шейман И.М. // Онтогенез.1981. Т.12. No.6. С.635Ч643.
8. Schaller H.Ch. // Cell Differ. 1976. V.5. No.1. P.1-11.
9. Шейман И.М., Тирас Х.П., Балобанова б.Ф. // Физиол. журн. СССР. 1989. Т.75. No.5. С.619-626.
10. Salo E., Baguna J. // J. Exp. Zool. 2002. V.292. P.528-539.