№ 8, 2000 г.
№ 8, 2000 г.
№ 8, 2000 г.
№ 8, 2000 г.
© Е.В. Райкова, Т.О. Напара, А.Ю. ИбрагимовЗагадочная паразитическая книдария
Е.В. Райкова, Т.О. Напара, А.Ю. Ибрагимов
Екатерина Викторовна Райкова, д.б.н.,
вед. н.с. лаборатории морфологии клетки Института цитологии РАН.
Татьяна Олеговна Напара, к.б.н., мл.н.с. той же лаборатории.
Алексей Юрьевич Ибрагимов, студент биофака ЛГУ.Книдарии, или стрекающие (Cnidaria), — древнейший из многоклеточных тип беспозвоночных животных. Надежный и важный систематический признак книдарий— стрекательные клетки (или книдоциты, или нематоциты, или крапивные клетки), которые служат им для ловли добычи, защиты, прикрепления к субстрату и передвижения. Книдарии, как правило, — свободноживущие формы, к ним относятся знакомые читателю гидра (широко распространенный пресноводный полип) и множество морских медуз. Объект же нашего внимания — настоящий гурман, живет не где-нибудь, а внутри икринок осетровых рыб! О существовании паразитических видов среди книдарий известно уже довольно давно. В морях обитают представители семейства наркомедуз (Narcomedusae, отряд Trachilida, класс Hydrozoa), которые паразитируют на стадии личинки в гастральной полости других медуз. Они прославились также тем, что во время эмбрионального развития один из бластомеров (клеток дробления яйца) становится клеткой-переносчиком (фороцитом) дробящегося зародыша, а потом участвует и в его питании [1].
В 1871 г. академик Ф.В.Овсянников нашел загадочного паразита (рис. 1) внутри икринок стерляди, который после вылупления в воде (рис. 2 и 3) приобретает вид гидроидного полипа2. Открытие внутриклеточного паразита (не из числа простейших, а из многоклеточных) стало настоящей сенсацией. Затем “овсянниковскую паразитную личинку” изучал О.А.Гримм, но он принял ее за личинку какого-то неведомого червя. И только в 1885 г. профессор М.М.Усов дал описание этого необычного животного, подтвердил его систематическую принадлежность к книдариям и назвал по сходству с гидрой и по манере передвижения Polypodium hydriforme.
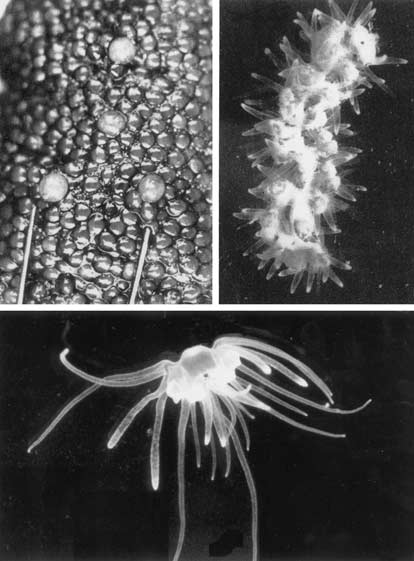 |
Рис. 1. Четыре зараженные икринки среди нормальной
зрелой икры стерляди (слева). Увел. 3.
Рис. 2. Столон после выхода из ооцита в воду. Увел. 6. Рис. 3. Свободноживущий полиподий с 24-мя щупальцами. Увел. 15. |
Этому организму вновь повезло, когда его исследованием основательно занялся московский зоолог А.Н. Липин. И тут сенсации последовали одна за другой. Самая главная из них состояла в том, что внутри стерляжьей икры полиподий вывернут наизнанку и имеет вид столона с почками, щупальца которых находятся внутри самих почек (рис. 4, стадия 1), а клетки энтодермы обращены к желтку стерляжьего яйца. Таким образом, полиподий как бы купается в окружающем его деликатесе. Перед нерестом стерляди полиподий снова выворачивается, но теперь как бы на лицевую сторону, втягивая при этом весь не съеденный желток в образующиеся гастральные полости каждой почки и благополучно вылупляется из икринки в воду (рис. 2 и 4, стадия 2). Как рачительный хозяин, он обеспечивает себя таким образом полноценной пищей на первое время свободной жизни [3].
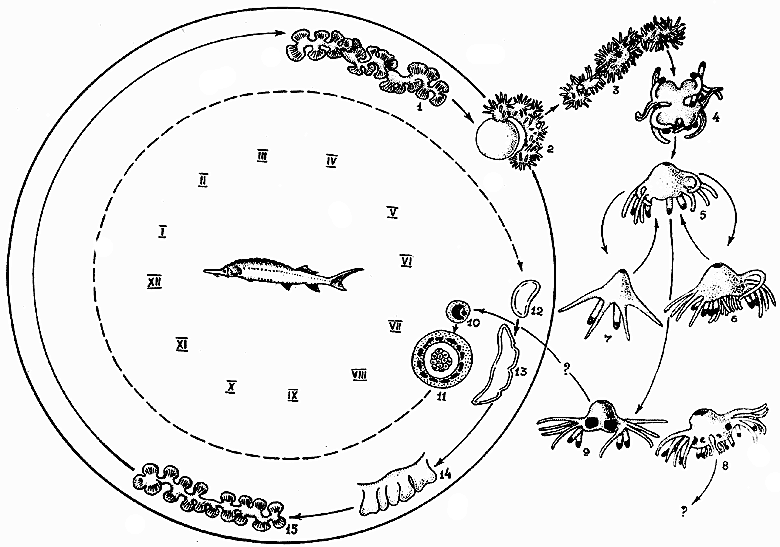
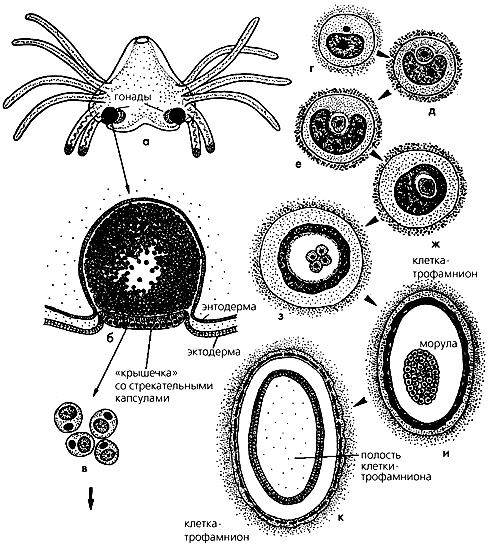
Рис. 4. Жизненный цикл Polypodium hydriforme. Паразитические стадии внутри круга, свободноживущие — снаружи. Римскими цифрами обозначены месяцы года, арабскими — стадии развития: 1 — зрелый столон с внутренними щупальцами в яйце перед икрометанием; 2 — выход из яйца столона с наружными щупальцами во время икрометания; 3 — свободный столон в воде; 4 — фрагмент столона; 5 — особь с 12 щупальцами; 6 — особь с 24 щупальцами; 7 — особь с шестью щупальцами; 8 — полиподий с 24 щупальцами и четырьмя “женскими” гонадами, роль которых неизвестна; 9 — особь с четырьмя “мужскими” гонадами (способ заражения ооцита неизвестен); 10 — двуядерная паразитическая клетка в молодом ооците; 11 — морула, окруженная трофамнионом; 12 — планула (трофамнион здесь и далее не показан); 13 — почкующаяся планула; 14 — столон без щупалец; 15 — столон с внутренними щупальцами.Выращивая свободноживущих полипов в аквариуме (рис. 3), Липин получил половозрелых особей с энтодермальными гонадами, снабженными выводными протоками, что уж совсем не укладывалось в характеристику класса Hydrozoa. Липин исследовал как женские, так и мужские гонады, но настоящих яиц и спермиев ему увидеть не удалось [4]. Что касается систематического положения, то Липин отнес Р.hydriforme к другому классу книдарий — Scyphozoa и в этом разошелся во мнениях с другим зоологом В.М. Шимкевичем, который придавал большое значение паразитическому образу жизни полиподия и считал его видом, близким к наркомедузам.
Впоследствии специалисты о полиподии если и вспоминали, то в основном как о зоологическом курьезе, причисляя его, как и губок, к Enantiozoa (вывернутым наизнанку животным). В учебниках зоологии и даже паразитологии о полиподии писали мелким шрифтом как о дополнении к отряду Hydrida, да и в спецкурсах о нем не упоминали.
Вновь заговорили об этом удивительном животном ихтиологи Ленинградского университета, которые вели работы с осетровыми на Волге и в других водоемах СССР. Оказалось, что Р.hydriforme серьезно угрожает запасам этих ценных рыб. Г.М.Персов, К.З.Трусов, москвички Т.А.Детлаф и А.С.Гинзбург нашли полиподия в икринках не только стерляди и даже не только у волжских рыб. Персов настоятельно советовал профессору В.А.Догелю, в то время заведующему кафедрой зоологии беспозвоночных Ленинградского университета, обратить внимание на эту проблему и исследовать полиподия. Догель сразу откликнулся:
“Особенно угрожающим является то обстоятельство, что Р.hydriforme есть паразит икры, т.е. органов размножения, а потому при сильной инвазии может привести к кастрации самок осетровых. Вот почему мы считаем своим долгом поднять сигнал тревоги и призвать к исследованию поднятого нами вопроса” [5].
Это было в 1940 г. А потом была война, затем послевоенная разруха. Ситуация с осетровыми действительно становилась тревожной. Персов говорил, что видел самок стерляди с тотальным заражением икры полиподием. В коллекции препаратов болезней рыб ГосНИОРХа хранится зафиксированный в формалине яичник такой несчастной стерляди из р.Камы, где нет практически ни одной здоровой икринки. Догель даже сравнил угрозу, нависшую со стороны полиподия осетровому стаду, с дамокловым мечом.
Под руководством Догеля в стенах ГосНИОРХа исследованием этой проблемы пришлось заняться и одному из авторов статьи (Е.В.Райковой). Основное внимание уделялось изучению годичного цикла и паразитических стадий Р.hydriforme, и новые неожиданные факты не заставили себя ждать. Оказалось, что планула, которую видели в ооцитах (икринках стерляди) и Усов, и Липин, также имеет обратное расположение зародышевых листков (рис. 4, стадии 12, 13). У всех книдарий (в том числе и у паразитических наркомедуз) планула — это стадия свободноплавающей личинки, которая обеспечивает расселение вида. Очень хотелось и нам найти планулу у выведенных из икринок полиподиев и проследить, как она заражает рыбу, как проникает в ооцит, как выворачивается наизнанку, чтобы поглощать клетками энтодермы желток стерляжьей икринки. Однако оказалось, что все эмбриональное развитие паразита вплоть до личинки происходит в мелких ооцитах младшей генерации, еще не имеющих желтка (рис. 4, стадии 10—12 и рис. 5, г—к), и стало быть никакой свободноплавающей планулы у полиподия просто не существует.
Рис. 5. Схема возникновения двуядерных клеток и преобразование двуядерной клетки в зародыш и трофамнион: а — свободноживущая особь с гонадами; б — гонада-гаметофор, содержащая зрелые двуядерные клетки; в — несколько одноклеточных двуядерных гамет при большом увеличении; г—к — паразитические стадии цикла внутри ооцитов осетровых; г — двуядерная клетка; д — вторая клеточная паразитическая стадия; е — малая клетка в углублении ядра большой клетки; ж — ядро большой клетки полностью окружает малую клетку; з — дробление малой клетки внутри большой, и — морула внутри полости большой клетки-трофамниона; к — планулообразная личинка внутри полости клетки-трофамниона.Эмбриогенез длится несколько лет (у остальных книдарий – всего несколько часов!) в соответствии с темпом развития ооцитов рыбы. Начальные стадии Р.hydriforme внутри ооцитов имеют вид клетки с двумя неравными ядрами (с микро- и макронуклеусом в одной цитоплазме) — прямо как у инфузорий (рис. 4, стадия 10; рис. 5, г). Все это казалось невероятным, однако и последующие события в эмбриональном развитии полиподия внутри икринок вызвали не меньшее удивление: двуядерная клетка делится и образует две неравные по размеру клетки, причем малая заключена внутри большой.
Стадия “клетка в клетке” (рис. 5, д—ж) длится до тех пор, пока ооциты не вступают в фазу большого роста.
Соответствие стадий развития паразита фазам оогенеза стерляди оказалось идеально строгим. Потом это было проверено и на других осетровых — белуге, шипе, осетре и севрюге. Таким образом, эмбриогенез у Р.hydriforme — самый медленный среди книдарий; клетка, из которой развивается зародыш, — самое мелкое и своеобразное яйцо, а стадия личинки — инвертированная и тоже самая длительная6.
Поскольку Р.hydriforme — метазойный внутриклеточный паразит, практически все проблемы, связанные с его изучением оказались в конечном счете цитологическими, а потому мы обратились за помощью к замечательному ученому-цитологу, профессору И.И.Соколову, возглавлявшему тогда лабораторию морфологии клетки в недавно созданном Институте цитологии АН СССР. Соколов был другом Догеля, настоящим исследователем-натуралистом, и никакие трудности объекта, никакие необычные, казавшиеся невозможными, данные не пугали его, а наоборот, вызывали только энтузиазм, который передавался его ученикам и сотрудникам.
Цитофотометрические исследования показали, что в системе “клетка в клетке” внутренняя малая (генеративная) клетка, как и ранние бластомеры (вплоть до 16-клеточной стадии), сохраняют количество ДНК, соответствующее гаплоидному набору хромосом. Это состояние генеративного ядра и ранних бластомеров длится у Р. hydriforme несколько лет. Диплоидность восстанавливается позже и пока невыясненным способом.
Все паразитические стадии (продукты деления малой клетки) развиваются в полости, образуемой большой клеткой с большим ядром (рис. 5, ж—к). Эта клетка-оболочка была названа трофамнионом по аналогии с трофической оболочкой вокруг зародыша у перепончатокрылых [7] (наездника — паразита гусениц).
Трофамнион Р.hydriforme — гомолог полярного тельца — полиплоидизируется, при этом концентрация ДНК увеличивается более чем в 500 раз, и в таком состоянии существует несколько лет, защищая и питая зародыш. Разрушается же он лишь незадолго до нереста рыбы. Выяснилось, что именно трофамнион, а не энтодермальные клетки столона, поглощает желток икринки.
Гистологическое исследование свободноживущих полиподиев также принесло интересные результаты. Подтвердились данные Липина о необычности гаметогенеза в гонадах полиподия. Оказалось, что у “мужских” особей (рис. 4, стадия 9; рис. 5,а) в гонадах (рис. 5,б) гаметогенез начинается, как типичный сперматогенез, а затем приобретает черты оогенеза, с двумя редукционными делениями. Во втором делении происходит только кариокинез (т.е. делится только ядро), а цитокинез блокируется и, вероятно, на многие годы. Клетка получается двуядерной, при этом одно ядро сразу же полиплоидизируется и становится макронуклеусом, а другое — остается гаплоидным (рис. 5,в). Гонада заполняется двуядерными клетками и закупоривается “крышечкой” (эктодермальной пластинкой, несущей стрекательные клетки) и превращается в гаметофор (рис. 5,б).
У “женских” особей (рис. 4, стадия 8) гонады — это сложный энтодермальный орган, состоящий из двух яичников с яйцеводами, открывающимися в гастральную полость. Ни мейотических делений, ни каких-либо характерных для яиц дейтоплазматических включений никому из исследователей наблюдать у половых клеток не удалось. Более того, женские гонады оказались нестабильными образованиями: они могут деградировать сразу после закладки, так и не развившись, могут разрушиться даже тогда, когда все их части уже полностью сформировались.
Удивительная находка была сделана ихтиологом И.И. Смольяновым. Исследуя развитие севрюги на Волге в районе Саратова, он нашел около десятка предличинок этой рыбы, на которые полиподии либо активно откладывали, либо уже отложили свои гаметофоры [8]. Смольянов, к его чести, не только определил незнакомые ему организмы, но и отправил тщательно зафиксированные пробы по нужному адресу для гистологического изучения. Побольше бы таких любознательных и настойчивых исследователей! На срезах удалось увидеть, как происходит процесс заражения рыб: стрекательные капсулы “крышечки” гаметофора выстреливают, при этом сама “крышечка” разрушается, а содержимое гаметофора (двуядерные половые клетки) вступает в контакт с покровами предличинки.
Благодаря этой находке мы убедились в правильности своих предположений и получили много новой информации.
Во-первых, исчезли сомнения в том, что половые клетки в гонадах культивируемых полиподиев вполне “нормальные” — они такие же двуядерные, как и в природе. Значит, гаметогенез действительно заканчивается образованием столь необычных двуядерных клеток.
Во-вторых, стало очевидным и предназначение гонад, выполняющих функцию гаметофоров.
В-третьих, найдено важное звено в цепи событий при заражении рыб: прикрепление гаметофоров полиподия происходит на стадии предличинок, когда покровы будущих осетров еще достаточно нежны.
Однако много еще неясного. Двуядерные клетки в гаметофорах и те, которые уже проникли внутрь ооцитов стерляди, различаются лишь по величине самих клеток и по концентрации ДНК в большом ядре (рис. 5,в и г). Малые ядра на обеих сравниваемых стадиях цикла оказались гаплоидными. Следовательно, развитие полиподия происходит без оплодотворения, т.е. партеногенетически, что очень редко встречается у книдарий, но характерно для других паразитов.
Возникла и очень пикантная цитологическая и общебиологическая проблема: развитие нового поколения паразита, заражающего самок осетровых, осуществляется за счет половых клеток, которые образовались в мужской гонаде! Спрашивается, для чего же тогда нужна гонада другого типа с выводными протоками и питающими клетками, которую мы считали женской?
Эта загадка не решена и поныне. Вероятно, в ходе эволюции под влиянием паразитического образа жизни у Р.hydriforme произошла переориентация пола мужских гонад. Женские гонады постепенно перестали давать зрелые яйца и сохранились как рудименты. Развитие пошло по типу партеногенеза на базе мужской гонады. Эта сугубо аберрантная черта в развитии Р.hydriforme — видоизмененный андрогенез [9], уникальное явление в животном мире, не описанное в естественных (не экспериментальных) условиях.
Находка Смольянова прибавила много и чисто паразитологических проблем. Если осетровые заражаются еще на стадии предличинки, то сколько же лет проходит, пока паразит займет свое место в ооците? Ведь осетровые растут медленно, и самая быстро созревающая из них стерлядь становится половозрелой лишь в 6—7 лет, а севрюга гораздо старше. Различает ли полип, откладывая гаметофоры на мальков, будущих самок или самцов? В последнем случае развитие, вероятно, приведет в тупик. Кроме того, судя по опыту культивирования и по находкам в природе, полипы созревают примерно к середине лета. В эту пору в Волге как раз и появляются вылупившиеся севрюжки, но ведь основной-то хозяин полиподия — стерлядь, а она нерестится всегда в конце мая, вместе с ее икринками выходят в воду и столоны полиподия (рис. 2). Предличинки стерляди вылупляются еще через неделю, когда столон полиподия в воде распадается до появления одиночных полипчиков (рис. 3; рис. 4, стадии 3—5), которые как раз успевают израсходовать весь запасенный желток и только-только начинают свою “книдарную” свободную жизнь, приступая к самостоятельной ловле добычи.
Жизнь свободноживущих Р.hydriforme также полна загадок. Немного странным кажется их облик. У них нет зонтика (рис. 3), но они умеют хорошо парить в воде, медленно передвигаться по дну, переставляя свои щупальца. Щупальца не окружают венчиком рот, как у других форм, а размещены по бокам, по шесть с каждой стороны: четыре тонких осязательных и два толстых опорных, на концах усаженных множеством стрекательных клеток (рис. 3).
Неясно, как полипы ориентируются, ведь у них не обнаружено никаких органов чувств, чем питаются в реке. Глотка у них тоже своеобразная — выстлана воротничковыми клетками, что очень характерно для малоподвижных или сидячих животных, например коралловых полипов, но только не для гидроидов.
Необычно и бесполое размножение Р.hydriforme. Паратомия, т.е. продольное деление, начинается всегда с аборального полюса, где закладываются новые щупальца, а ротовой конус разделяется в последнюю очередь, что другим книдариям не свойственно. Образовавшиеся после деления две сестринские особи получают с одной стороны старый комплект щупалец, а с другой, со стороны борозды деления, — шесть новых. В связи с этим полиподии всегда в той или иной степени асимметричны (рис. 3). Основная форма — 12-щупальцевые особи (рис. 4, стадия 5), но если деление запаздывает, появляются 24-щупальцевые (рис. 4, стадия 6), а если новые щупальца почему-либо не закладываются, что обычно бывает перед гибелью, то получаются 6-щупальцевые полипы (рис. 4, стадия 7).
Таким образом, перечисленные аберрантные черты организации полиподия явно не укладываются в характеристику класса Hydrozoa. В то же время его паразитический образ жизни и черты эмбрионального развития, принцип архитектоники столона и свободноживущих форм, гомология осей тела, строение щупалец, а также одинаковый, по классификации Р.Вейла [10], тип стрекательных капсул свидетельствуют в пользу родства полиподия с наркомедузами. И наконец, как мы уже отмечали, у Р.hydriforme ярко выражены “протозойные” черты, сходные с организацией как инфузорий, так и миксоспоридий (Myxozoa), которых раньше относили к простейшим.
В последние годы много внимания уделяется филогенетическим отношениям миксоспоридий и книдарий. Еще в 1938 г. Вейл предположил, что миксоспоридии — это редуцированные книдарии [11]. Более полувека спустя М.Э.Сиддал с коллегами, изучив молекулярные характеристики последовательностей 18S РНК миксоспоридий и полиподия (использовался и присланный нами Р.hydriforme), убедились в том, что это — сестринские группы [12]. Подтвердилось это и в большой работе по изучению филогении многоклеточных животных, основанной на анализе 18S рибосомной ДНК [13]. В результате же других молекулярных исследований, в которых миксоспоридий сравнивали с книдариями (но не с полиподием), выявлено сходство миксоспоридий с двусторонне-симметричными животными [14] (Bilateria).
Действительно, из-за паразитического образа жизни у Р.hydriforme много общего с миксоспоридиями. Изложение этой проблемы заслуживает однако специальной статьи, которая уже готовится к печати. Мы стремились показать, что, хотя во всех современных учебниках полиподий и считается наркомедузой, у него достаточно своих аберрантных черт, выводящих его даже за рамки класса гидроидных (Hydrozoa). Радиально симметричные стрекательные клетки у полиподия весьма своеобразны. Книдоциль (чувствительный волосок) расположен над крышечкой ядовитой капсулы, а не рядом с ней, как у остальных книдарий; стрекательная нить у большинства капсул снабжена шипами [15] (по принятой классификации Вейла они рассматриваются как капсулы с гладкой нитью).
В мезоглее (студенистом веществе между экто- и эндодермой) полиподия обнаружены нехарактерные для гидроидов структуры: внеклеточные микротрубочки [16], амебоциты (бесцветные подвижные клетки, защищающие организм от инородных частиц, подобно лейкоцитам позвоночных животных), которые не долговечны и быстро гибнут; а также нервные элементы [17]. Кроме того, мышечная система полиподия, и только у него одного, представлена лишь самостоятельными гладкими мускульными клетками, лежащими в мезоглее [18].
Итак, какое же положение должен занять Р.hydriforme в системе типа Cnidaria в настоящее время? Правильно установить систематическую принадлежность животного так же важно, как определить место химического элемента в системе Менделеева. Базируясь в основном на данных собственных исследований и сравнивая их с литературными, мы согласны с тем, что P.hydriforme и Myxozoa — это сестринские группы. На наш взгляд, они вместе с паразитическими наркомедузами могут представлять собой отдельный таксон паразитических книдарий в ранге класса, эволюция которого под влиянием паразитического образа жизни шла по пути упрощения.
* * *
Нерешенных проблем еще очень много, остаются невыясненными многие загадки на стыке фундаментальных исследований по цитологии, паразитологии, эмбриологии, эволюционной морфологии.
За последние двадцать лет P.hydriforme был обнаружен в икринках осетровых рыб Нового Света. Очень бы хотелось сравнить “наших старосветских” полиподиев с их заокеанскими собратьями, тем более что виды осетровых там совсем другие, а паразиты в их икре морфологически неотличимы. Заодно неплохо бы разобраться и с определением видов у наших собственных полиподиев из разных водоемов и разных хозяев — стерляди, осетра, севрюги, шипа и белуги, обитающих в бассейнах Каспия, Амура, Арала, в сибирских реках. Мы понимаем, что нужно торопиться, ибо численность осетровых быстро сокращается и отнюдь не по вине полиподия, а от загрязнения наших морей и рек, хищнического вылова и вопиющей бесхозяйственности. Уже сейчас в Астрахани не всегда увидишь осетра, а тем более белугу, и найти подходящих рыб для сбора полиподия становится все труднее. Но мы полны энтузиазма, и нам бы очень хотелось разгадать тайну P.hydriforme как можно скорее.
Работа выполнена при поддержке Российского фонда фундаментальных исследований. Грант № 97-04-48988.
ЛИТЕРАТУРА
1 Hyman L. The invertevrates. N. Y.; London, 1940. V.1.
2 Овсянников Ф.В. // Тр. 3-го съезда рус. естествоиспытателей. Киев, 1873. С.351—358.
3 Lipin A.N. // Uss. Zool. Jahrb. (Anat.). 1911. Bd. 31. S.317—426.
4 Lipin A.N. // Zool. Jahrb. (Anat.). 1925. Bd.47. S.541—635.
5 Догель В.А. // Зоол. журн. 1940. Т.19. №2. С.321—323.
6 Raikova E.V. // J. Parasitol. 1994. V.80. P.1—22.
7 Raikova E.V. // Cell and Tissue Res. 1980. V.206. P.487—500; Иванова-Казас О.М. Очерки по сравнительной эмбриологии перепончатокрылых. Л., 1961.
8 Смольянов И.И., Райкова Е.В. // Докл. АН СССР. 1961.T.141. С.1271—1274.
9 Райкова Е.В. // Цитология. 1985. T.27. №4. C.391—401.
10 Weill R. // Trav. stat. Zool. Wimereux. 1934. V.10—11. P.1—701.
11 Weill R. // Trav. Stat. Zool. Wime€reaux. 1938. V.13. P.727—744.
12 Siddall M.E., Martin D.S., Bridge D. et al. // J. Parasitol. 1995. V.81. P.961—967.
13 Zrzavy J., Mihulka S., Kepka P. et al. // Cladistics. 1998. V.14. P.249—285.
14 Smothers J.F., Dohlen C.D. von, Smith L.H.Jr., Spall R.D. // Science. 1994. V.265. P.1719—1721; Schlegel M., Lom J., Stechmann A. et al. // Arch. Protistenkunde. 1996. Bd.147. S.1—9.
15 Raikova E.V. // Zoologica scripta. 1990. V.19. P.1—11; Ибрагимов А.Ю. // Цитология. 1999. T.41. C.200—209.
16 Raikova E.V., Napara T.O. // 7-th Workshop on Hydroid development. Evangelische Akademie Tutzing. 22—25 Sept. 1997. P.99.
17 Napara T.O. // 7-th Workshop on Hydroid development. Evangelische Akademie Tutzing. 22—25 Sept. 1997. P.98.
18 Райкова Е.В., Напара Т.О. // Цитология. T.41. №5. C.425—431.
 Июль 2000 |