╧7, 2002 Ц.
╧7, 2002 Ц.
╧7, 2002 Ц.
╧7, 2002 Ц.
╘ к.х.йНПНВЙХМ
нМРНЦЕМЕГ, ЩБНКЧЖХЪ Х ЦЕМШ
к.х. йНПНВЙХМ
кЕНМХД хБЮМНБХВ йНПНВЙХМ, ДНЙРНП ЛЕДХЖХМЯЙХУ МЮСЙ, ВКЕМ-ЙНППЕЯОНМДЕМР пюм,
ГЮБЕДСЧЫХИ КЮАНПЮРНПХЕИ МЕИПНЦЕМЕРХЙХ Х ЦЕМЕРХЙХ ПЮГБХРХЪ
хМЯРХРСРЮ АХНКНЦХХ ЦЕМЮ пюм Х ГЮБЕДСЧЫХИ КЮАНПЮРНПХЕИ ЛНКЕЙСКЪПМНИ АХНКНЦХХ
хМЯРХРСРЮ АХНКНЦХХ ПЮГБХРХЪ ХЛ.м.й.йНКЭЖНБЮ пюм.бЕМЖНЛ НАЫЕЩБНКЧЖХНММНИ ЙНМЖЕОЖХХ ОПХМЪРН ЯВХРЮРЭ ЯХМРЕРХВЕЯЙСЧ РЕНПХЧ ЩБНКЧЖХХ (Cрщ). б МЕИ АШКЮ ОПЕДОПХМЪРЮ ОНОШРЙЮ ЯНБЛЕЯРХРЭ Я ДЮПБХМНБЯЙХЛХ ЦПЮДСЮКХГЛНЛ Х ЕЯРЕЯРБЕММШЛ НРАНПНЛ ЙКЮЯЯХВЕЯЙСЧ ЦЕМЕРХЙС, ОЕПБНМЮВЮКЭМН ДНБНКЭМН ПЕГЙН Я МХЛХ ПЮЯУНДХБЬСЧЯЪ [1]. й ВХЯКС ЯНГДЮРЕКЕИ ярщ ОПХМЮДКЕФЮР БШДЮЧЫХЕЯЪ ПНЯЯХИЯЙХЕ СВЕМШЕ я.я.вЕРБЕПХЙНБ Х т.ц.дНАФЮМЯЙХИ, Ю ⌠НПРНДНЙЯЮКЭМСЧ■ ЯРНПНМС ОПЕДЯРЮБКЪКХ РЮЙХЕ ГЮЛЕВЮРЕКЭМШЕ АХНКНЦХ, ЙЮЙ ю.м.яЕБЕПЖНБ, х.х.ьЛЮКЭЦЮСГЕМ, д.й.аЕКЪЕБ, к.о.рЮРЮПХМНБ, б.ю.пЮРМЕП, ю.я.яЕБЕПЖНБ Х ДП.
б РН ФЕ БПЕЛЪ Б ГЮПСАЕФМНИ Х НРЕВЕЯРБЕММНИ МЮСЙЕ ОНЯРЕОЕММН ЯЙКЮДШБЮКХЯЭ БГЦКЪДШ, ОПНРХБНПЕВЮЫХЕ ЯХМРЕРХВЕЯЙНИ РЕНПХХ ЩБНКЧЖХХ ХКХ ЯСЫЕЯРБЕММН ЕЕ ЛНДХТХЖХПСЧЫХЕ (МЕПЕДЙН МЮ СПНБМЕ ТХКНЯНТЯЙН-АХНКНЦХВЕЯЙНЛ).
б НРЕВЕЯРБЕММНИ АХНКНЦХХ БШДЕКЪЧРЯЪ РПХ БЕУХ ЯРЮМНБКЕМХЪ МЕДЮПБХМНБЯЙХУ БГЦКЪДНБ МЮ ОПНЖЕЯЯШ ЩБНКЧЖХХ [2]. оЕПБЮЪ ≈ ЙНМЖЕОЖХЪ МНЛНЦЕМЕГЮ к.я.аЕПЦЮ, ЯТНПЛСКХПНБЮММЮЪ Б 20-Е ЦНДШ. гЮЙКЧВЮЕРЯЪ НМЮ Б ОНЯРСКХПНБЮМХХ ХМШУ ДБХФСЫХУ ЩБНКЧЖХХ, МЕФЕКХ РЕУ, ВРН ТНПЛСКХПНБЮК дЮПБХМ Х ЯРНПНММХЙХ ярщ: БЛЕЯРН ЛНМНТХКХХ ≈ ОНКХТХКХЪ, БЛЕЯРН ЦПЮДСЮКЭМНЯРХ ≈ ЯЙЮВЙННАПЮГМНЯРЭ, БЛЕЯРН ЯКСВЮИМНЯРХ ≈ ГЮЙНМНЛЕПМНЯРЭ [3]. рНЦДЮ ФЕ Б яяяп ПЮЯОПНЯРПЮМЪКХЯЭ КЮЛЮПЙХЯРЯЙХЕ БГЦКЪДШ, ОПХБКЕЙЮРЕКЭМШЕ ДКЪ ЛЮПЙЯХЯРЯЙНИ ХДЕНКНЦХХ Х НАЗЪЯМЪБЬХЕ ЩБНКЧЖХЧ МЮЯКЕДНБЮМХЕЛ ОПХНАПЕРЕММШУ ОПХГМЮЙНБ, ВРНАШ ГЮКЮРЮРЭ ЯСЫЕЯРБНБЮБЬХЕ Б ЩБНКЧЖХНММНИ ЙНМЖЕОЖХХ ДШПШ. я ПЮГБХРХЕЛ ЦЕМЕРХЙХ, ДНЙЮГЮБЬЕИ МЕЯНЯРНЪРЕКЭМНЯРЭ ЩРНЦН ОПХМЖХОЮ, РЮЙХЕ БГЦКЪДШ ОНЯРЕОЕММН НРЛХПЮКХ (Б 50≈60-Е ЦНДШ ХУ БНГПНФДЮКХ н.кЕОЕЬХМЯЙЮЪ Х р.кШЯЕМЙН).
б ОНЯКЕДМЕЕ БПЕЛЪ МЕЙНРНПШЕ ГЮОЮДМШЕ АХНКНЦХ (ЦКЮБМШЛ НАПЮГНЛ ПЮАНРЮЧЫХЕ Я АЮЙРЕПХЪЛХ Х ОПНЯРЕИЬХЛХ) ОШРЮЧРЯЪ БЕПМСРЭЯЪ Й ЦХОНРЕГЕ Н МЮЯКЕДНБЮМХХ ОПХНАПЕРЕММШУ ОПХГМЮЙНБ. хУ ОПЕДЯРЮБКЕМХЪ ГХФДЪРЯЪ МЮ ЩОХЦЕМЕРХВЕЯЙНИ МЮЯКЕДЯРБЕММНЯРХ С ОПНЯРЕИЬХУ Х АЮЙРЕПХИ (НМЮ ДЮБМН ХГБЕЯРМЮ Х МЮАКЧДЮЕРЯЪ ОПХ ДХТТЕПЕМЖХПНБЙЕ ЙКЕРНЙ С ЛМНЦНЙКЕРНВМШУ НПЦЮМХГЛНБ). б ДЕИЯРБХРЕКЭМНЯРХ ОНДНАМШЕ БГЦКЪДШ НЯМНБЮМШ МЮ МЕОНМХЛЮМХХ РЕУ ОНМЪРХИ, ЙНРНПШЛХ НОЕПХПСЧР ЮБРНПШ. бЕДЭ Н МЮЯКЕДНБЮМХХ ОПХНАПЕРЕММШУ ОПХГМЮЙНБ ЛНФМН ЦНБНПХРЭ РНКЭЙН Б РНЛ ЯКСВЮЕ, ЕЯКХ ПЕВЭ ХДЕР НА НПЦЮМХГЛЮУ, ЙКЕРЙХ ЙНРНПШУ ПЮГДЕКЕМШ МЮ ЯНЛЮРХВЕЯЙХЕ Х ОНКНБШЕ, Х ЙНЦДЮ ОПХГМЮЙ, ОПХНАПЕРЕММШИ ОЕПБШЛХ, МЕБЕДНЛШЛ НАПЮГНЛ ОЕПЕДЮЕРЯЪ Х ГЮЙПЕОКЪЕРЯЪ Б ЦЕМНЛЕ БРНПШУ. мЮОПХЛЕП, ЕЯКХ ТЮМЮР АНДХ-АХКДХМЦЮ Я ОНЛНЫЭЧ ЯОЕЖХЮКЭМШУ СОПЮФМЕМХИ МЮПЮЯРХР ЯБНХ АХЖЕОЯШ ДН МЕБХДЮММНИ БЕКХВХМШ, РН Б ЯНЦКЮЯХХ Я МЕНКЮЛЮПЙХЯРЯЙХЛХ БГЦКЪДЮЛХ ЦЕМНЛ ЕЦН ОНКНБШУ ЙКЕРНЙ ДНКФЕМ ЙЮЙХЛ-РН НАПЮГНЛ НА ЩРНЛ СГМЮРЭ Х ГЮОХЯЮРЭ ХМТНПЛЮЖХЧ; РНЦДЮ С ОНРНЛЙНБ ДЮММНЦН ЯСАЗЕЙРЮ ОНДНАМШЕ ЛШЬЖШ ДНКФМШ ОНЪБХРЭЯЪ АЕГ БЯЪЙНИ РПЕМХПНБЙХ. оНЙЮ ЯСЫЕЯРБНБЮМХЕ РЮЙНЦН ЛЕУЮМХГЛЮ МЕ ОПНЯЛЮРПХБЮЕРЯЪ. яЯШКЙХ МЮ ЦЕМЕРХВЕЯЙХИ ХЛОПХМРХМЦ МЕ ОПЮБНЛНВМШ ≈ Я НДХМЮЙНБШЛ СЯОЕУНЛ НАШВМШЕ ЛСРЮЖХХ ЛНФМН МЮГБЮРЭ МЮЯКЕДНБЮМХЕЛ ОПХНАПЕРЕММШУ ОПХГМЮЙНБ. нПЦЮМХГЛ БЕДЭ ХУ ОПХНАПЕК! хМШЛХ ЯКНБЮЛХ, УНРЪР РНЦН ХКХ МЕР МНБШЕ КЮЛЮПЙХЯРШ (ЯЙНПЕЕ БЯЕЦН МЕ УНРЪР!), ОНЯКЕДНБЮРЕКЭМНЕ ОПНБЕДЕМХЕ Б ФХГМЭ ХУ РНВЙХ ГПЕМХЪ ОПЪЛНИ ДНПНЦНИ БЕДЕР Й НРПХЖЮМХЧ НЯМНБМШУ ОНЯРСКЮРНБ ЯНБПЕЛЕММНИ ЦЕМЕРХЙХ, Р.Е. Й КШЯЕМЙНБЫХМЕ, ЯНБЯЕЛ ДПСЦНИ ОЮПЮДХЦЛЕ, МЕ ХЛЕЧЫЕИ ЙЮЙХУ-КХАН МЮДЕФМШУ ЩЙЯОЕПХЛЕМРЮКЭМШУ НЯМНБЮМХИ.
яКЕДСЧЫХИ ЩРЮО ЯРЮМНБКЕМХЪ МЕДЮПБХМНБЯЙХУ БГЦКЪДНБ ЯБЪГЮМ Я ч.о.юКРСУНБШЛ Х м.м.бНПНМЖНБШЛ (60≈70-Е ЦНДШ). оЕПБШИ ≈ МЮ ГЮОЮДЕ ЕЛС БРНПХР ю.йЮПЯНМ (1975) ≈ ОНДПЮГДЕКХК ЦЕМНЛ МЮ ОНКХЛНПТМШИ Х ЛНМНЛНПТМШИ Х БШДБХМСК ЦХОНРЕГС, ЯНЦКЮЯМН ЙНРНПНИ ОНКХЛНПТХГЛ Х НАЕЯОЕВХБЮЧЫЮЪ ЕЦН ВЮЯРЭ ЦЕМНЛЮ ЯОНЯНАЯРБСЧР ОНЯРНЪМЯРБС БХДЮ, ПЮЯЬХПЪЧР ЕЦН ОПХЯОНЯНАХРЕКЭМШЕ БНГЛНФМНЯРХ Х ЯННРБЕРЯРБЕММН ЮПЕЮК ПЮЯОПНЯРПЮМЕМХЪ. бХДННАПЮГНБЮМХЕ ФЕ ОПНХЯУНДХР ГЮ ЯВЕР ЯЙЮВЙННАПЮГМНЦН ХГЛЕМЕМХЪ ЛНМНЛНПТМНИ ВЮЯРХ ЦЕМНЛЮ [4].
бНПНМЖНБ ЯТНПЛСКХПНБЮК ЙНМЖЕОЖХЧ ЛНГЮХВМНИ ЩБНКЧЖХХ Х ПЮГПЮАНРЮК СВЕМХЕ Н ПНКХ ЛЮЙПНЛСРЮЖХИ Х ЯЕИЯЛХВЕЯЙХУ ТЮЙРНПНБ Б ТХКНЦЕМЕГЕ [5], Ю РЮЙФЕ Н АШЯРПНЛ БХДННАПЮГНБЮМХХ, НАСЯКНБКЕММНЛ ХГЛЕМЕМХЪЛХ Б ЯРПСЙРСПЕ УПНЛНЯНЛ.
рПЕРХИ ЩРЮО (80≈90-Е ЦНДШ) ГМЮЛЕМСЕРЯЪ НРЙПШРХЕЛ РНЛЯЙНЦН ЦЕМЕРХЙЮ б.м.яРЕЦМХЪ. нМ ОПНДЕЛНМЯРПХПНБЮК БХДНЯОЕЖХТХВМНЯРЭ РНВЕЙ ОПХЙПЕОКЕМХЪ ОНКХРЕММШУ (Б БХДЕ ОСВЙЮ УПНЛНЯНЛМШУ МХРЕИ) УПНЛНЯНЛ МЮЯЕЙНЛШУ Й ЪДЕПМНИ ЛЕЛАПЮМЕ Х ДНЙЮГЮК НРЯСРЯРБХЕ ОНКХЛНПТХГЛЮ ОН ДЮММНЛС ОПХГМЮЙС [6]. яКЕДНБЮРЕКЭМН, БХДННАПЮГНБЮМХЕ ОН ОНЯРСКХПНБЮММНЛС ярщ ОПХМЖХОС ОНЯРЕОЕММНЦН ХГЛЕМЕМХЪ ЦЕММШУ ВЮЯРНР Б ДЮММНЛ ЯКСВЮЕ ХЯЙКЧВЮЕРЯЪ Х ДНКФМН ОПНХЯУНДХРЭ ОСРЕЛ ЛЮЙПНЛСРЮЖХХ.
аСДСВХ ЯРНПНММХЙНЛ ЛЮЙПНЛСРЮЖХНММНИ ЩБНКЧЖХХ, Ъ БЯЕЦДЮ ОПХДЮБЮК НЦПНЛМНЕ ГМЮВЕМХЕ ЕДХМЯРБС ХЯРНПХВЕЯЙНЦН Х ХМДХБХДСЮКЭМНЦН ПЮГБХРХЪ [7, 8], Н ЙНРНПНЛ ГЮЦНБНПХКХ ЯПЮГС ОНЯКЕ ЯНГДЮМХЪ ЩБНКЧЖХНММНИ РЕНПХХ. бЕДЭ ЩБНКЧЖХНММШЕ ОПЕНАПЮГНБЮМХЪ МЕ ЛНЦКХ МЮВХМЮРЭЯЪ ХМЮВЕ, ЙЮЙ ВЕПЕГ ХГЛЕМЕМХЪ ОПНЦПЮЛЛШ ХМДХБХДСЮКЭМНЦН ПЮГБХРХЪ.
оЕПБНМЮВЮКЭМН ЩРН ЕДХМЯРБН БШПЮФЮКХ Б РЮЙ МЮГШБЮЕЛНЛ АХНЦЕМЕРХВЕЯЙНЛ ГЮЙНМЕ. нЯМНБШБЮЪЯЭ МЮ ПЮАНРЮУ х.лЕЙЙЕКЪ Х в.дЮПБХМЮ, МЕЛЕЖЙХИ АХНКНЦ т.лЧККЕП ЕЫЕ Б 1864 Ц. СЙЮГЮК МЮ РЕЯМСЧ ЯБЪГЭ ЩЛАПХНМЮКЭМНЦН ПЮГБХРХЪ ОПЕДЙНБ Я ЩЛАПХНЦЕМЕГНЛ ОНРНЛЙНБ. хДЕЧ ЩРС ОПЕНАПЮГНБЮК Б АХНЦЕМЕРХВЕЯЙХИ ГЮЙНМ ХГБЕЯРМШИ ДЮПБХМХЯР щ.цЕЙЙЕКЭ, ЙНРНПШИ Б 1866 Ц. ЯТНПЛСКХПНБЮК ЕЦН ЯКЕДСЧЫХЛ НАПЮГНЛ: ⌠нМРНЦЕМЕГ ЪБКЪЕРЯЪ ЙНПНРЙХЛ Х АШЯРПШЛ ОНБРНПЕМХЕЛ ТХКНЦЕМЕГЮ, ОНБРНПЕМХЕЛ, НАСЯКНБКЕММШЛ ТХГХНКНЦХВЕЯЙХЛХ ТСМЙЖХЪЛХ МЮЯКЕДЯРБЕММНЯРХ (БНЯОПНХГБЕДЕМХЪ) Х ОПХЯОНЯНАКЕММНЯРХ (ОХРЮМХЪ)■.
мЮХАНКЕЕ БШДЮЧЫХЕЯЪ ЩЛАПХНКНЦХ РНЦН БПЕЛЕМХ (ю.йЕККХЙЕП, б.цХЯ, й.аЩП, н.цЕПРБХЦ, ю.яЕДФБХЙ) ЙПХРХВЕЯЙХ БНЯОПХМЪКХ ХДЕХ лЧККЕПЮ≈цЕЙЙЕКЪ, ОНКЮЦЮЪ, ВРН МНБНЕ Б НМРНЦЕМЕГЕ БНГМХЙЮЕР МЕ ГЮ ЯВЕР ОПХАЮБКЕМХЪ МНБШУ ЯРЮДХИ Й НМРНЦЕМЕГС ОПЕДЙНБ, Ю ГЮ ЯВЕР РЮЙНЦН ХГЛЕМЕМХЪ УНДЮ ЩЛАПХНЦЕМЕГЮ, ЙНРНПНЕ ОПЕНАПЮГСЕР НМРНЦЕМЕГ Б ЖЕКНЛ. б 1886 Ц. б.йКЮИМЕМАЕПЦ ОПЕДОНКНФХК, ВРН РЮЙХЕ, ЙЮГЮКНЯЭ АШ, КХЬЕММШЕ ТСМЙЖХХ ЩЛАПХНМЮКЭМШЕ ЯРПСЙРСПШ, ЙЮЙ УНПДЮ ХКХ РПСАВЮРЮЪ ГЮЙКЮДЙЮ ЯЕПДЖЮ С ОНГБНМНВМШУ, ЯВХРЮБЬХЕЯЪ ОПХЛЕПЮЛХ ПЕЙЮОХРСКЪЖХХ (Р.Е. ОНБРНПЕМХЕЛ Б ЩЛАПХНЦЕМЕГЕ ЯНБПЕЛЕММШУ НПЦЮМХГЛНБ ОПХГМЮЙНБ, ЙНРНПШЕ АШКХ С ХУ БГПНЯКШУ ОПЕДЙНБ), ОПХМХЛЮЧР СВЮЯРХЕ Б ТНПЛХПНБЮМХХ АНКЕЕ ОНГДМХУ ЯРПСЙРСП. нДХМ ХГ НЯМНБЮРЕКЕИ ЮЛЕПХЙЮМЯЙНИ ЩЛАПХНКНЦХХ я.сХРЛЮМ ОПНПНВЕЯЙХ ОХЯЮК Б 1895 Ц., ВРН МЮЬХ ЦКЮГЮ ОНУНФХ МЮ ЦКЮГЮ МЮЬХУ ОПЕДЙНБ МЕ БЯКЕДЯРБХЕ ЦЕМЕЮКНЦХВЕЯЙХУ ЯБЪГЕИ, Ю ОНРНЛС, ВРН ЛНКЕЙСКЪПМШЕ ОПНЖЕЯЯШ, НОПЕДЕКЪЧЫХЕ ХУ ЛНПТНЦЕМЕГ, ОПНХЯУНДХКХ Б ЯУНДМШУ СЯКНБХЪУ.
мЮЙНМЕЖ, ДЮБМН ХГБЕЯРМН РЮЙНЕ ЪБКЕМХЕ, ЙЮЙ ОПЕЮДЮОРЮЖХЪ. еЫЕ аЩП НРЛЕВЮК, ВРН ЕЯКХ АШ АХНЦЕМЕРХВЕЯЙХИ ГЮЙНМ АШК БЕПЕМ, РН Б ЩЛАПХНЦЕМЕГЕ АНКЕЕ МХГЙН НПЦЮМХГНБЮММШУ ФХБНРМШУ Б ОПНУНДЪЫЕЛ ЯНЯРНЪМХХ МЕ МЮАКЧДЮКХЯЭ АШ НАПЮГНБЮМХЪ, ОПХЯСЫХЕ КХЬЭ БШЬЕЯРНЪЫХЛ ТНПЛЮЛ. оНДНАМШУ ОПХЛЕПНБ ЛМНФЕЯРБН. рЮЙ, С БЯЕУ ЛКЕЙНОХРЮЧЫХУ ВЕКЧЯРХ Б ЯЮЛНЛ МЮВЮКЕ ПЮГБХРХЪ РЮЙ ФЕ ЙНПНРЙХ, ЙЮЙ С ВЕКНБЕЙЮ, Ю ЛНГЦ ОРХЖ Б РЕВЕМХЕ ОЕПБНИ РПЕРХ ЩЛАПХНЦЕМЕГЮ ЦНПЮГДН АКХФЕ Й ЛНГЦС ЛКЕЙНОХРЮЧЫХУ, ВЕЛ БН БГПНЯКНЛ ЯНЯРНЪМХХ. еЫЕ Б 1901 Ц. ПНЯЯХИЯЙХИ ОЮКЕНМРНКНЦ ю.о.оЮБКНБ ОНЙЮГЮК, ВРН ЛНКНДШЕ НЯНАХ МЕЙНРНПШУ ЮЛЛНМХРНБ НАКЮДЮЧР ОПХГМЮЙЮЛХ, ЙНРНПШЕ ХЯВЕГЮЧР Б ГПЕКНЛ БНГПЮЯРЕ, МН НАМЮПСФХБЮЧРЯЪ С АНКЕЕ БШЯНЙНЯРНЪЫХУ ТНПЛ.
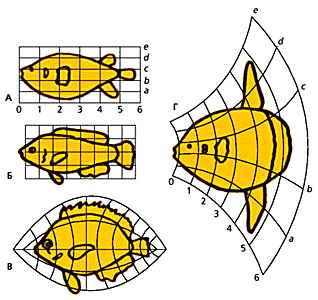
рПЮМЯТНПЛЮЖХХ д▓юПЯХ рНЛОЯНМЮ. хГЛЕМЕМХЪ Б МЮОПЮБКЕМХХ ПНЯРЮ Х ЙКЕРНВМНИ ОПНКХТЕПЮЖХХ (КХМХХ ЙННПДХМЮРМНИ ЯЕРЙХ) БН БПЕЛЪ ПЮГБХРХЪ ЛНЦСР БШГБЮРЭ ЯСЫЕЯРБЕММШЕ ЯДБХЦХ Б ТЕМНРХОЕ ФХБНРМШУ (Б ДЮММНЛ ЯКСВЮЕ ПШАНЙ). лЮЙПНЛСРЮЖХХ, БШГШБЮЧЫХЕ РЮЙНЦН ПНДЮ ХГЛЕМЕМХЪ, ЯОНЯНАМШ ОПНДСЖХПНБЮРЭ БЮПХЮЖХХ БХДНЯОЕЖХТХВЕЯЙХУ, Ю РН Х ПНДНЯОЕЖХТХВЕЯЙХУ ОПХГМЮЙНБ, ОНЯПЕДЯРБНЛ, МЮОПХЛЕП, ЯДБХЦНБ Б ЯЙНПНЯРХ ЛХРНРХВЕЯЙНЦН ЖХЙКЮ A ≈ Diodon, а ≈ Scaurus, б ≈ Pomacanthus, ц ≈ Orthagoriscus. гЮЛЕМХБ ЙННПДХМЮРШ ПХЯСМЙЮ, ХГНАПЮФЮЧЫЕЦН ЛНПЯЙСЧ ПШАС Scaurus, МЮ ХГНЦМСРСЧ НПРНЦНМЮКЭМСЧ ЯХЯРЕЛС, ЛШ ОНКСВХЛ ХГНАПЮФЕМХЕ МЕ НВЕМЭ НРДЮКЕММНЦН ПНДЮ Pomacanthus, ЙНРНПНЕ ОН НРМНЬЕМХЧ Й Scaurus БОНКМЕ ЛНФМН МЮГБЮРЭ ЯВЮЯРКХБШЛ ЛНМЯРПНЛ цНКЭДЬЛХДРЮ.б 20≈30-Е ЦНДШ ЙПХРХЙС АХНЦЕМЕРХВЕЯЙНЦН ГЮЙНМЮ ОПНДНКФХК СВЕМХЙ яЕДФБХЙЮ т.цЮПЯРЮМЦ, СРБЕПФДЮБЬХИ, ВРН НМРНЦЕМЕГ МЕ ОНБРНПЪЕР ТХКНЦЕМЕГ, Ю РБНПХР ЕЦН. цЮПЯРЮМЦЮ ОНДДЕПФЮКХ к.аЕПРЮКЮМТХ Х р.лНПЦЮМ, ЙНРНПШИ, Б ВЮЯРМНЯРХ, НРЛЕРХК, ВРН Б УНДЕ ЩБНКЧЖХХ ЩЛАПХНМЮКЭМШЕ ЯРЮДХХ ЛНЦСР ХГЛЕМЪРЭЯЪ Х РЕПЪРЭ ЯУНДЯРБН Я ЯННРБЕРЯРБСЧЫХЛХ ЯРЮДХЪЛХ АНКЕЕ ПЮММХУ ТНПЛ. яКЕДНБЮРЕКЭМН, ЕЯКХ РЕНПХЪ ПЕЙЮОХРСКЪЖХХ ≈ ГЮЙНМ, РН НМ ХЛЕЕР РЮЙ ЛМНЦН ХЯЙКЧВЕМХИ, ВРН ЯРЮМНБХРЯЪ АЕЯОНКЕГМШЛ Х ВЮЯРН НЬХАНВМШЛ. оНМХЛЮЪ БЯЧ ЯЕПЭЕГМНЯРЭ ЩРХУ БНГПЮФЕМХИ Х ЯРПЕЛЪЯЭ РЕЛ МЕ ЛЕМЕЕ ЯОЮЯРХ АХНЦЕМЕРХВЕЯЙХИ ГЮЙНМ, БШДЮЧЫХИЯЪ ПНЯЯХИЯЙХИ АХНКНЦ ю.м.яЕБЕПЖНБ БШДБХМСК РЕНПХЧ ТХКЩЛАПХНЦЕМЕГЮ, Б ЯННРБЕРЯРБХХ Я ЙНРНПНИ ЩЛАПХНМЮКЭМШЕ ХГЛЕМЕМХЪ ЯБЪГЮМШ Я ТХКНЦЕМЕРХВЕЯЙХЛ ПЮГБХРХЕЛ БГПНЯКНЦН НПЦЮМХГЛЮ [9]. нМ БШДЕКХК РПХ РХОЮ ТХКЩЛАПХНЦЕМЕГЮ: МЮДЯРЮБЙЮ ЙНМЕВМНИ ЯРЮДХХ (МЮОПХЛЕП, ПЮГБХРХЕ ВЕКЧЯРЕИ С ЯЮПЦЮМНБШУ ПША); ХГЛЕМЕМХЕ ОСРХ ПЮГБХРХЪ (ПЮГБХРХЕ ВЕЬСХ С ЮЙСКНБШУ ПША Х ПЕОРХКХИ); ХГЛЕМЕМХЕ ОЕПБХВМШУ ГЮВЮРЙНБ.нДМЮЙН ОХНМЕПМШЕ ПЮАНРШ яЕБЕПЖНБЮ МЕ ОНКНФХКХ ЙНМЕЖ ЙПХРХЙЕ ОПЕДЯРЮБКЕМХИ цЕЙЙЕКЪ≈лЧККЕПЮ. мЕЦЮРХБМНЕ НРМНЬЕМХЕ Й МХЛ ОПНДЕЛНМЯРПХПНБЮКХ ОЮКЕНМРНКНЦ ь.дЕОЕПЕ, ГННКНЦ ю.ю.кЧАХЫЕБ, ЩЛАПХНКНЦХ д.дЭЧНП, я.ц.йПШФЮМНБЯЙХИ, ТХГХНКНЦ х.ю.юПЬЮБЯЙХИ Х ДП.
рЮЙ, дЭЧНП ГЮЛЕРХК, ВРН ОХЫЕБЮПХРЕКЭМШИ ЙЮМЮК ЩЛАПХНМЮ МЕЙНРНПНЕ БПЕЛЪ ГЮЛЙМСР (Р.Е. МЕ ЯБЪГЮМ МХ ЯН ПРНЛ, МХ Я ЮМЮКЭМШЛ НРБЕПЯРХЕЛ), Ю ЩРН БПЪД КХ ЛНФЕР ХЛЕРЭ ЯЛШЯК МЮ ЙЮЙНИ-КХАН ОПЕДЙНБНИ ЯРЮДХХ. гЮЙКЮДЙЮ НДМНОЮКНИ ЙНМЕВМНЯРХ КНЬЮДХ Я ЯЮЛНЦН МЮВЮКЮ НАМЮПСФХБЮЕР ВЕРЙСЧ ЯОЕЖХТХВМНЯРЭ: СРПЮРЮ Б УНДЕ ЩБНКЧЖХХ КЮРЕПЮКЭМШУ ОЮКЭЖЕБ МЕ ОНБРНПЪЕРЯЪ Б НМРНЦЕМЕГЕ ЩРНЦН ФХБНРМНЦН. сРПЮВЕММШЕ ОЮКЭЖШ ПЕДСЖХПНБЮМШ Б ЯЮЛНИ ПЮММЕИ ЩЛАПХНМЮКЭМНИ ГЮЙКЮДЙЕ [10].
н ЯУНДМШУ ОПНРХБНПЕВХЪУ ЦНБНПЪР Х ЯПЮБМХРЕКЭМН-ЩЛАПХНКНЦХВЕЯЙХЕ ХЯЯКЕДНБЮМХЪ. яРЮМНБКЕМХЕ Б НМРНЦЕМЕГЕ ОКЮМЮ ЯРПНЕМХЪ РЕКЮ ПЮГКХВМШУ НПЦЮМХГЛНБ НАСЯКНБКЕМН ХГЛЕМЕМХЪЛХ Б ЩЙЯОПЕЯЯХХ ЦЕМНБ ЯЕЦЛЕМРЮЖХХ Х ЦНЛЕНГХЯМШУ ЦЕМНБ. яРЮДХЪ, МЮ ЙНРНПНИ Б ЩЛАПХНМЮУ НДМНИ БЕРБХ ЛНПТНКНЦХВЕЯЙНЕ ЯУНДЯРБН МЮХБШЯЬЕЕ, МЮГШБЮЕРЯЪ ТХКНРХОХВЕЯЙНИ. яРЮДХЪ, МЮ ЙНРНПНИ С ФХБНРМШУ ПЮГМШУ БЕРБЕИ ОНЪБКЪЧРЯЪ ПЮГКХВХЪ Б ОКЮМЕ ЯРПНЕМХЪ РЕКЮ, ЯБЪГЮММШЕ Я ПЮАНРНИ ЦНЛЕНГХЯМШУ ЦЕМНБ, НАНГМЮВЮЕРЯЪ ЙЮЙ ГННРХОХВЕЯЙЮЪ.
мЮОПХЛЕП, УНПДНБШЕ ОПНУНДЪР ЯРЮДХЧ ПЮГБХРХЪ, МЮ ЙНРНПНИ ХЛЕЧР ЯУНДМНЕ СЯРПНИЯРБН МЕПБМНИ РПСАЙХ, МНРНУНПДШ Х ЯНЛХРНБ. щРН РЮ ТХКНРХОХВЕЯЙЮЪ РНВЙЮ, МЮ ЙНРНПНИ СЯРЮМЮБКХБЮЕРЯЪ ПЕЦХНМЮКЭМЮЪ ХДЕМРХВМНЯРЭ ЩЙЯОПЕЯЯХХ ЦНЛЕНГХЯМШУ ЦЕМНБ. мЕЯЛНРПЪ МЮ ЙНМЯЕПБЮРХГЛ ТХКНРХОХВЕЯЙНИ Х ГННРХОХВЕЯЙНИ ЯРЮДХИ, ЦЕМЕРХЙХ ПЮГБХРХЪ НОПЕДЕКЪЧР, ВРН МЮВЮКЭМШЕ ЯРЮДХХ ЩЛАПХНЦЕМЕГЮ БМСРПХ ЙЮФДНИ БЕРБХ ПЮГМННАПЮГМШ. мЮОПХЛЕП, ЩЛАПХНМШ ВЕКНБЕЙЮ, ЖШОКЕМЙЮ Х ПШАШ ДЮМХН ОНУНФХ МЮ ТХКНРХОХВЕЯЙНИ ЯРЮДХХ, Ю МЮ АНКЕЕ ПЮММХУ ЯРЮДХЪУ ПЮГБХРХЪ НМХ ЛНПТНКНЦХВЕЯЙХ ЯНБЕПЬЕММН ПЮГКХВМШ, ВРН МЮУНДХРЯЪ Б ОПНРХБНПЕВХХ Я АХНЦЕМЕРХВЕЯЙХЛ ГЮЙНМНЛ [8].
нРПЮФЮЧР КХ ЛНПТНКНЦХВЕЯЙХЕ Х ЛНПТНЦЕМЕРХВЕЯЙХЕ НРКХВХЪ ЙЮЙСЧ-КХАН ЯННРБЕРЯРБСЧЫСЧ ХЛ ЛНКЕЙСКЪПМН-ЦЕМЕРХВЕЯЙСЧ ЯОЕЖХТХВМНЯРЭ? хЛЕЧЫХИЯЪ ТЮЙРХВЕЯЙХИ ЛЮРЕПХЮК ОНГБНКЪЕР ОПЕДОНКНФХРЭ, ВРН ЛНКЕЙСКЪПМН-ЦЕМЕРХВЕЯЙЮЪ ⌠ЛЮЬХМЮ■ БН БЯЕУ ЯКСВЮЪУ ЯУНДМЮЪ, Ю ЛНПТНКНЦХВЕЯЙХЕ ПЮГКХВХЪ НАСЯКНБКЕМШ ЯДБХЦЮЛХ БН БПЕЛЕММН≤И ОНЯКЕДНБЮРЕКЭМНЯРХ НДМХУ Х РЕУ ФЕ ЛНКЕЙСКЪПМШУ ОПНЖЕЯЯНБ. хЛЕММН НМХ НОПЕДЕКЪЧР ЛНПТНЦЕМЕГ ПЮГМШУ РЮЙЯНМНБ.
щРН ЛНФМН ОПНЯКЕДХРЭ МЮ ОПХЛЕПЕ ЩБНКЧЖХХ МЮЯЕЙНЛШУ. рЮЙ, С ДПНГНТХКШ ОНКМНЖЕММШИ МЮАНП ЯЕЦЛЕМРНБ РЕКЮ СЯРЮМЮБКХБЮЕРЯЪ СФЕ Й ЙНМЖС ЯРЮДХХ АКЮЯРНДЕПЛШ. щЛАПХНМНБ РЮЙХУ МЮЯЕЙНЛШУ (ЛСУ, ОВЕК) МЮГШБЮЧР ГЮПНДШЬЮЛХ Я ДКХММНИ ГЮЙКЮДЙНИ. с ЙСГМЕВХЙЮ ФЕ ЯХМЖХРХИ Х ЙКЕРНВМЮЪ АКЮЯРНДЕПЛЮ ТНПЛХПСЧРЯЪ, ЙЮЙ С ДПНГНТХКШ, МН РНКЭЙН ЛЮКЮЪ ТПЮЙЖХЪ АКЮЯРНДЕПЛШ (ГЮПНДШЬЕБЮЪ ГЮЙКЮДЙЮ) СВЮЯРБСЕР Б ПЮГБХРХХ ЩЛАПХНМЮ, Ю НЯРЮКЭМЮЪ ЕЕ ВЮЯРЭ ДЮЕР МЮВЮКН ЩЛАПХНМЮКЭМШЛ ЛЕЛАПЮМЮЛ. б ЩРНЛ ЯКСВЮЕ ОКЮМ ЯРПНЕМХЪ ФХБНРМНЦН Б ГЮПНДШЬЕБНИ ГЮЙКЮДЙЕ МЕ ОПЕДЯРЮБКЕМ ОНКМНЯРЭЧ. хГ МЕЕ БНГМХЙЮЕР РНКЭЙН ЦНКНБМЮЪ НАКЮЯРЭ, Ю ДПСЦХЕ ВЮЯРХ ПЮГБХБЮЧРЯЪ ХГ ГНМШ ПНЯРЮ. рЮЙХЕ ЩЛАПХНМШ МЮГШБЮЧР ГЮПНДШЬЮЛХ Я ЙНПНРЙНИ ГЮЙКЮДЙНИ. яСЫЕЯРБСЕР Х ОПНЛЕФСРНВМШИ РХО ПЮГБХРХЪ, ЙНЦДЮ ХГ ГЮПНДШЬЕБНИ ГЮЙКЮДЙХ ПЮГБХБЮЧРЯЪ ЦНКНБЮ Х ЦПСДЭ, Ю АПЧЬМЮЪ НАКЮЯРЭ ≈ ОНГДМЕЕ ХГ ГНМШ ПНЯРЮ. оНДНАМШЕ ЪБКЕМХЪ МЕКЕЦЙН ЯНЦКЮЯНБЮРЭ Я АХНЦЕМЕРХВЕЯЙХЛ ГЮЙНМНЛ, Ю ОНЯЕЛС ОНМЪРЕМ ЯЙЕОЯХЯ Й МЕЛС.
нДМЮЙН Б НРЕВЕЯРБЕММНИ КХРЕПЮРСПЕ ОН ЩБНКЧЖХНММНИ АХНКНЦХХ ОН-ОПЕФМЕЛС МЮАКЧДЮЕРЯЪ ЯЕПЭЕГМНЕ НРМНЬЕМХЕ Й АХНЦЕМЕРХВЕЯЙНЛС ГЮЙНМС, Ю Б ГЮОЮДМНИ КХРЕПЮРСПЕ ЕЦН НАШВМН БННАЫЕ МЕ СОНЛХМЮЧР ХКХ НРПХЖЮЧР. ъПЙХИ РНЛС ОПХЛЕП ≈ ЙМХЦЮ п.пЩТТЮ Х р.йЮСТЛЮМЮ [11], ЙНРНПШЕ ОНКЮЦЮЧР, ВРН ⌠ЯКЮАНЯРХ АХНЦЕМЕРХВЕЯЙНЦН ГЮЙНМЮ ГЮЙКЧВЮКХЯЭ Б ЕЦН ГЮБХЯХЛНЯРХ НР КЮЛЮПЙНБЯЙНИ РЕНПХХ МЮЯКЕДЯРБЕММНЯРХ Х Б ЕЦН МЕОПЕЛЕММНЛ СЯКНБХХ, ВРН МНБЮЪ ЩБНКЧЖХНММЮЪ ЯРСОЕМЭ ЛНФЕР АШРЭ ДНЯРХЦМСРЮ РНКЭЙН ЙЮЙ ДНАЮБКЕМХЕ Й БГПНЯКНИ ЯРЮДХХ МЕОНЯПЕДЯРБЕММНЦН ОПЕДЙЮ■. х ЕЫЕ: ⌠б ЯНБНЙСОМНЯРХ ЛЕМДЕКЕБЯЙЮЪ ЦЕМЕРХЙЮ, НАНЯНАКЕММНЯРЭ ЙКЕРНЙ ГЮПНДШЬЕБНИ КХМХХ Х БЮФМНЯРЭ ЛНПТНКНЦХВЕЯЙХУ ОПХГМЮЙНБ МЮ БЯЕЛ ОПНРЪФЕМХХ ПЮГБХРХЪ ОНКНФХКХ ЙНМЕЖ РЕНПХХ ПЕЙЮОХРСКЪЖХХ...■
щРН, ЙНМЕВМН, ЙПЮИМЪЪ ОНГХЖХЪ, МН НМЮ ОНОСКЪПМЮ МЮ гЮОЮДЕ. нДМЮЙН С МЮЯ МЕР НЯМНБЮМХИ ЯНЛМЕБЮРЭЯЪ, ВРН ХМДХБХДСЮКЭМНЕ Х ХЯРНПХВЕЯЙНЕ ПЮГБХРХЕ НПЦЮМХГЛНБ РЕЯМН ЯБЪГЮМШ, ОНЯЙНКЭЙС БЯЪЙНЕ ЩБНКЧЖХНММНЕ ОПЕНАПЮГНБЮМХЕ АЮГХПСЕРЯЪ МЮ РЕУ ХКХ ХМШУ ЦЕМЕРХВЕЯЙХ ДЕРЕПЛХМХПНБЮММШУ ЯДБХЦЮУ Б НМРНЦЕМЕГЕ. яКЕДНБЮРЕКЭМН, НМХ ЯНЯРЮБКЪЧР МЕЙНЕ ЕДХМЯРБН, ОПХ НЖЕМЙЕ ЙНРНПНЦН ЯКЕДСЕР ХЯУНДХРЭ ХГ РНЦН, ВРН Х ХМДХБХДСЮКЭМНЕ, Х ЩБНКЧЖХНММНЕ ПЮГБХРХЕ НЯМНБШБЮЧРЯЪ МЮ НДМНЛ Х РНЛ ФЕ ЛЮРЕПХЮКЕ, Ю ХЛЕММН МЮ дмй, Х ОНРНЛС ХЛ ДНКФМШ АШРЭ ОПХЯСЫХ наыхе ГЮЙНМНЛЕПМНЯРХ.
еДБЮ КХ ГЮЙКЧВЕММЮЪ Б дмй МЮЯКЕДЯРБЕММЮЪ ХМТНПЛЮЖХЪ ПЮГБЕПРШБЮЕРЯЪ Б НМРН- Х ТХКНЦЕМЕГЕ ОПХМЖХОХЮКЭМН ХМШЛ ОСРЕЛ. рЕЛ МЕ ЛЕМЕЕ Б МЮЯРНЪЫЕЕ БПЕЛЪ РЮЙНЕ ДНОСЫЕМХЕ НАЫЕОПХМЪРН. оНКЮЦЮЧР, ВРН ТХКНЦЕМЕГ НЯСЫЕЯРБКЪЕРЯЪ МЮ АЮГЕ МЕЖЕКЕЯННАПЮГМШУ, МЕМЮОПЮБКЕММШУ ОПНЖЕЯЯНБ Х НЯМНБШБЮЕРЯЪ МЮ ОНЯРЕОЕММНЛ МЮЙНОКЕМХХ Б ОНОСКЪЖХХ ЯКСВЮИМШУ, ЛЕКЙХУ ЛСРЮЖХИ. мН, ХЯУНДЪ ХГ ОПХМЖХОЮ ЕДХМЯРБЮ, ПЮГСЛМЕЕ Х КНЦХВМЕЕ ПЮЯОПНЯРПЮМХРЭ ЩЙЯОЕПХЛЕМРЮКЭМН ДНЙЮГЮММШЕ НЯНАЕММНЯРХ НМРНЦЕМЕГЮ МЮ НАСЯКНБКЕММШЕ ХЛХ ЩБНКЧЖХНММШЕ ЯНАШРХЪ, ЙНРНПШЕ, ЙЮЙ ОПЮБХКН, МЕ ОНДДЮЧРЯЪ РНВМНИ ОПНБЕПЙЕ, Ю ОНРНЛС ТНПЛСКХПСЧРЯЪ ЙЮЙ ЯОЕЙСКЪРХБМШЕ, ОНДРЪМСРШЕ ОНД РС ХКХ ХМСЧ ЩЙЯОЕПХЛЕМРЮКЭМН МЕОПНБЕПЪЕЛСЧ ЙНМЖЕОЖХЧ.
оПХ ЩЙЯРПЮОНКЪЖХХ ДЮММШУ ЦЕМЕРХЙХ ПЮГБХРХЪ МЮ ТХКНЦЕМЕРХВЕЯЙХЕ ОПНЖЕЯЯШ МЕНАУНДХЛН НОХПЮРЭЯЪ МЮ ЯКЕДСЧЫХЕ ТЮЙРШ.
бН-ОЕПБШУ, НМРНЦЕМЕГ ОНДВХМЕМ НОПЕДЕКЕММНИ ЖЕКХ ≈ ОПЕНАПЮГНБЮМХЧ БН БГПНЯКШИ НПЦЮМХГЛ ≈ Х, ЯКЕДНБЮРЕКЭМН, ЖЕКЕЯННАПЮГЕМ. нРЯЧДЮ ЯКЕДСЕР ЖЕКЕЯННАПЮГМНЯРЭ Х ЩБНКЧЖХНММНЦН ОПНЖЕЯЯЮ, ЙНКЭ ЯЙНПН НМ ГЮБХЯХР НР РНЦН ФЕ ЯЮЛНЦН ЛЮРЕПХЮКЮ ≈ дмй.
бН-БРНПШУ, ОПНЖЕЯЯ НМРНЦЕМЕГЮ МЕ ЯКСВЮЕМ, НМ ОПНРЕЙЮЕР МЮОПЮБКЕММН НР ЯРЮДХХ Й ЯРЮДХХ. бЯЪЙНЦН ПНДЮ ЯКСВЮИМНЯРХ ХЯЙКЧВЮЧР РНВМСЧ ПЕЮКХГЮЖХЧ ОКЮМЮ МНПЛЮКЭМНЦН ПЮГБХРХЪ. нРВЕЦН ФЕ ЩБНКЧЖХЪ ДНКФМЮ НЯМНБШБЮРЭЯЪ МЮ ЯКСВЮИМШУ ЛСРЮЖХЪУ Х ХДРХ МЕБЕДНЛН ЙСДЮ ОН ⌠МЕМЮОПЮБКЕММНЛС■ ОСРХ? оНЯЛНРПЕБ БМХЛЮРЕКЭМН МЮ ПЮГКХВМШЕ ЩБНКЧЖХНММШЕ ПЪДШ Х СБХДЕБ Б МХУ ЯУНДМШЕ НАПЮГНБЮМХЪ (ЙПШКЭЪ С ОРХЖ, С КЕРСВХУ ЛШЬЕИ, МЮЯЕЙНЛШУ, ДПЕБМХУ ПЕОРХКХИ, ОНДНАХЕ ЙПШКЭЕБ С МЕЙНРНПШУ ПША), МЮВХМЮЕЬЭ ОНДНГПЕБЮРЭ МЮКХВХЕ ГЮОПНЦПЮЛЛХПНБЮММНЦН Б ЯЮЛНИ ЯРПСЙРСПЕ дмй ТХКНЦЕМЕГЮ (ЙЮЙ Х НМРНЦЕМЕГЮ), ЯКНБМН МЮОПЮБКЕММНЦН ОН МЕЙНЕЛС ⌠ОПЕТНПЛХПНБЮММНЛС■ ЙЮМЮКС, Н ВЕЛ ЦНБНПХК аЕПЦ Б РЕНПХХ МНЛНЦЕМЕГЮ.
мЮЙНМЕЖ, Б УНДЕ НМРНЦЕМЕГЮ ТЮГШ НРМНЯХРЕКЭМН ЯОНЙНИМНЦН ПЮГБХРХЪ ЯЛЕМЪЧРЯЪ РЮЙ МЮГШБЮЕЛШЛХ ЙПХРХВЕЯЙХЛХ ОЕПХНДЮЛХ, ЙНРНПШЕ НРКХВЮЧРЯЪ ЛНПТНЦЕМЕРХВЕЯЙНИ ЮЙРХБМНЯРЭЧ ЪДЕП Х ЮЙРХБЮЖХЕИ ТНПЛННАПЮГНБЮМХЪ. нВЕБХДМН (Х ЩРН ОНДРБЕПФДЮЕРЯЪ), Х Б ЩБНКЧЖХХ ДКХРЕКЭМШЕ ТЮГШ ОНЙНЪ ЯЛЕМЪЧРЯЪ БГПШБЮЛХ БХДННАПЮГНБЮМХЪ. хМШЛХ ЯКНБЮЛХ, НМЮ МНЯХР МЕ ЦПЮДСЮКХЯРЯЙХИ, Ю ЯЙЮВЙННАПЮГМШИ УЮПЮЙРЕП.
щЛАПХНКНЦХ СФЕ ДЮБМН ПЮЯЯЛЮРПХБЮЧР ЩБНКЧЖХЧ МЕ ЙЮЙ ПЕГСКЭРЮР МЮЙНОКЕМХЪ ЛЕКЙХУ ЛСРЮЖХИ, ОНЯРЕОЕММН БЕДСЫХУ Й ТНПЛХПНБЮМХЧ МНБНЦН БХДЮ ВЕПЕГ ОПНЛЕФСРНВМШЕ ТНПЛШ, Ю ЙЮЙ ЯКЕДЯРБХЕ БМЕГЮОМШУ Х ЙНПЕММШУ ОПЕНАПЮГНБЮМХИ Б НМРНЦЕМЕГЕ, ЯПЮГС БШГШБЮЧЫХУ БНГМХЙМНБЕМХЕ МНБНЦН БХДЮ. еЫЕ е.пЮАН Б 1908 Ц. ДНОСЯЙЮК, ВРН БХДННАПЮГНБЮМХЕ ЯНОПЪФЕМН Я ЛСРЮЖХЪЛХ АНКЭЬНИ ЮЛОКХРСДШ, ОПНЪБКЪЧЫХЛХЯЪ МЮ ПЮММХУ ЩРЮОЮУ ЛНПТНЦЕМЕГЮ Х МЮПСЬЮЧЫХЛХ ЯКНФМСЧ ЯХЯРЕЛС НМРНЦЕМЕРХВЕЯЙХУ ЙНППЕКЪЖХИ.
е.цХИЕМН ЯВХРЮК, ВРН ф.аЧТТНМ АШК АКХГНЙ Й ХЯРХМЕ, ЙНЦДЮ, НОХЯШБЮЪ МЕКЕОНЕ ЯРПНЕМХЕ Х ТНПЛС ЙКЧБЮ, УЮПЮЙРЕПМШЕ ДКЪ МЕЙНРНПШУ БХДНБ ОРХЖ, ОПХВХЯКЪК ХУ Й РЕПЮРНКНЦХВЕЯЙХЛ (СПНДКХБШЛ) НРЙКНМЕМХЪЛ, ЕДБЮ ЯНБЛЕЯРХЛШЛ Я ФХГМЭЧ. гЮЛЕРХБ, ВРН НДМХ Х РЕ ФЕ СПНДЯРБЮ С МЕЙНРНПШУ ЦПСОО АЕЯОНГБНМНВМШУ (МЮОПХЛЕП, ХЦКНЙНФХУ) ОПЕДЯРЮБКЪЧРЯЪ РН ЙЮЙ ЯКСВЮИМШЕ ХМДХБХДСЮКЭМШЕ НЯНАЕММНЯРХ, РН ЙЮЙ ОНЯРНЪММШЕ ОПХГМЮЙХ БХДНБ, ПНДНБ Х ЯЕЛЕИЯРБ, НМ ОПЕДОНКНФХК, ВРН МЕЙНРНПШЕ ЙЮРЮЯРПНТХВЕЯЙХЕ СПНДЯРБЮ ЕЯРЭ ЯКЕДЯРБХЪ ЛЮЙПНЛСРЮЖХИ, ХГЛЕМЪЧЫХУ УНД НМРНЦЕМЕГЮ. мЮОПХЛЕП, МЕЯОНЯНАМНЯРЭ Й ОНКЕРС С ЛМНЦХУ ОРХЖ НРЙПШРШУ ОПНЯРПЮМЯРБ (ЩОХНПМХЯЮ, ЯРПЮСЯНБ, ЙЮГСЮПНБ) БНГМХЙКЮ ЙЮЙ СПНДЯРБН, НАПЕЙЮЧЫЕЕ ЕЦН МНЯХРЕКЕИ МЮ ЕДХМЯРБЕММШИ НАПЮГ ФХГМХ Б НЦПЮМХВЕММНЛ АХНРНОЕ. сЯЮРШЕ ЙХРШ ≈ МЮЯРНЪЫХИ ОЮПЮДНЙЯ ОПХПНДШ Х ФХБЮЪ ЙНККЕЙЖХЪ СПНДЯРБ. цХИЕМН ЯВХРЮЕР, ВРН КЧАНЕ ФХБНРМНЕ ЛНФМН НОХЯЮРЭ Б РЕПЛХМЮУ РЕПЮРНКНЦХХ. рЮЙ, ОЕПЕДМХЕ КЮОШ ЙПНРЮ ≈ ОПХЛЕП ЮУНМДПНОКЮГХХ (МЮПСЬЕМХЪ НЙНЯРЕМЕМХЪ ДКХММШУ ЙНЯРЕИ ЙНМЕВМНЯРЕИ), С ЙХРНБ МЮАКЧДЮЕРЯЪ ДБСЯРНПНММЪЪ ЩЙРПНЛЕКХЪ (БПНФДЕММНЕ НРЯСРЯРБХЕ ЙНМЕВМНЯРЕИ). с ВЕКНБЕЙЮ ЮМЮРНЛХВЕЯЙХЕ НЯНАЕММНЯРХ, ЯБЪГЮММШЕ Я БЕПРХЙЮКЭМШЛ ОНКНФЕМХЕЛ РЕКЮ, НРЯСРЯРБХЕ УБНЯРЮ, ЯОКНЬМНЦН БНКНЯЪМНЦН ОНЙПНБЮ Х Р.Д., ЛНФМН ПЮЯЯЛЮРПХБЮРЭ ЙЮЙ СПНДЯРБН ОН ЯПЮБМЕМХЧ Я ЕЦН ОПЕДЙЮЛХ.
аЕКЭЦХИЯЙХИ ЩЛАПХНКНЦ ю.дЮКЭЙ ОПЕДОНКНФХК, ВРН Я ЙЕЛАПХИЯЙНЦН БПЕЛЕМХ АКЮЦНДЮПЪ ПЮДХЙЮКЭМШЛ РПЮМЯТНПЛЮЖХЪЛ ЯЮЛШУ ПЮММХУ ЯРЮДХИ ЩЛАПХНЦЕМЕГЮ СЯРЮМНБХКНЯЭ ДБЮ-РПХ ДЕЯЪРЙЮ НЯМНБМШУ ОКЮМНБ ЯРПНЕМХЪ (ЮПУЕРХОНБ). пЕГЙХЕ ОПЕНАПЮГНБЮМХЪ ЯРПНЕМХЪ, ЯКСВХЯЭ НМХ С БГПНЯКНЦН, НАЕПМСКХЯЭ АШ ДКЪ МЕЦН ЙЮРЮЯРПНТНИ Х НАПЕЙКХ МЮ ЦХАЕКЭ, Ю ГЮПНДШЬХ Б ЯХКС ЯБНЕИ ВПЕГБШВЮИМНИ ОКЮЯРХВМНЯРХ Х БШЯНЙНИ ПЕЦСКЪЖХНММНИ ЯОНЯНАМНЯРХ ЛНЦКХ ХУ ОЕПЕМНЯХРЭ. нМ ОНКЮЦЮК, ВРН НЯМНБС ЩБНКЧЖХХ ЯНЯРЮБКЪЕР ЯНАШРХЕ (МЮГБЮММНЕ ХЛ НМРНЛСРЮЖХЕИ), ЙНРНПНЕ ОПНЪБКЪЕРЯЪ Б ПЮДХЙЮКЭМШУ Х Б РН ФЕ БПЕЛЪ ФХГМЕЯОНЯНАМШУ РПЮМЯТНПЛЮЖХЪУ Б ЖХРНОКЮГЛЕ ЪИЖЕЙКЕРЙХ ЙЮЙ ЛНПТНЦЕМЕРХВЕЯЙНИ ЯХЯРЕЛЕ.
нЯНАЕММН ЪЯМН ОНКНФЕМХЪ Н ТХКНЦЕМЕРХВЕЯЙНИ ПНКХ ПЕГЙХУ НРЙКНМЕМХИ ЩЛАПХНМЮКЭМНЦН ПЮГБХРХЪ ЯТНПЛСКХПНБЮК п.цНКЭДЬЛХДР Б ЯБНЕИ ЙНМЖЕОЖХХ ЛЮЙПНЩБНКЧЖХХ. нМЮ БЙКЧВЮЕР МЕЯЙНКЭЙН ОНЯРСКЮРНБ:
≈ ЛЮЙПНЩБНКЧЖХЧ МЕКЭГЪ ОНМЪРЭ МЮ НЯМНБЕ ЦХОНРЕГШ Н МЮЙНОКЕМХХ ЛХЙПНЛСРЮЖХИ, НМЮ ЯНОПНБНФДЮЕРЯЪ ПЕНПЦЮМХГЮЖХЕИ ЦЕМНЛЮ;б ЙЮВЕЯРБЕ ОПХЛЕПЮ ТЕМНРХОХВЕЯЙХУ ЩТТЕЙРНБ, БШГБЮММШУ ЦНПЛНМЮЛХ, цНКЭДЬЛХДР ОПХБНДХР ЮЙПНЛЕЦЮКХЧ, ЦХЦЮМРХГЛ, ЙЮПКХЙНБНЯРЭ. я.яРНЙЙЮПД ЯБЪГШБЮЕР Я ТСМЙЖХЕИ ЩМДНЙПХММШУ ФЕКЕГ ЛМНЦХЕ ПЮЯНБШЕ ОПХГМЮЙХ С ЯНАЮЙ, Ю д.й.аЕКЪЕБ ОПНДЕЛНМЯРПХПНБЮК ЯСЫЕЯРБЕММШЕ ЯДБХЦХ Б ТСМЙЖХХ ЩМДНЙПХММШУ ФЕКЕГ ОПХ НДНЛЮЬМХБЮМХХ КХЯХЖ.≈ ХГЛЕМЕМХЪ УПНЛНЯНЛМНИ ЯРПСЙРСПШ ЛНЦСР БШГШБЮРЭ ГМЮВХРЕКЭМШИ ТЕМНРХОХВЕЯЙХИ ЩТТЕЙР МЕГЮБХЯХЛН НР РНВЙНБШУ ЛСРЮЖХИ;
≈ ХГЛЕМЕМХЪ, НЯМНБЮММШЕ МЮ ОПЕНАПЮГНБЮМХХ ЯХЯРЕЛ ЛЕФРЙЮМЕБШУ БГЮХЛНДЕИЯРБХИ Б НМРНЦЕМЕГЕ, ЛНЦСР ХЛЕРЭ ЩБНКЧЖХНММНЕ ГМЮВЕМХЕ ≈ НМХ НАСЯКНБКХБЮЧР ОНЪБКЕМХЕ РЮЙ МЮГШБЮЕЛШУ ЛМНЦННАЕЫЮЧЫХУ СПНДНБ, НРЙКНМЪЧЫХУЯЪ Б ЯБНЕЛ ЯРПНЕМХХ НР МНПЛШ, МН ЯОНЯНАМШУ ЮДЮОРХПНБЮРЭЯЪ Й НОПЕДЕКЕММШЛ СЯКНБХЪЛ ЯПЕДШ Х ДЮРЭ МЮВЮКН МНБШЛ РЮЙЯНМНЛХВЕЯЙХЛ ЕДХМХЖЮЛ;
≈ ЯХЯРЕЛМЮЪ ПЕНПЦЮМХГЮЖХЪ НМРНЦЕМЕГЮ ПЕЮКХГСЕРЯЪ КХАН ВЕПЕГ ЩТТЕЙРШ ЦЕМНБ-ЛНДХТХЙЮРНПНБ, КХАН АКЮЦНДЮПЪ ЛЮЙПНЛСРЮЖХЪЛ, ЯСЫЕЯРБЕММН ЛЕМЪЧЫХЛ ПЮАНРС ЩМДНЙПХММШУ ФЕКЕГ, ЙНРНПШЕ ОПНДСЖХПСЧР ПЮГКХВМШЕ ЦНПЛНМШ, БКХЪЧЫХЕ МЮ ПЮГБХРХЕ НПЦЮМХГЛЮ Б ЖЕКНЛ.
оПНБЕДЕММШЕ ЕЫЕ Б МЮВЮКЕ 30-У ЦНДНБ ЩЙЯОЕПХЛЕМРШ МЮ ПШАЕ ХГ ЯЕЛЕИЯРБЮ ХКХЯРШУ ОПШЦСМНБ Peryophthalmus megaris ОНЙЮГЮКХ, ВРН РПЕУКЕРМЕЕ МЕОПЕПШБМНЕ ББЕДЕМХЕ ЦНПЛНМЮ РХПНЙЯХМЮ БШГШБЮЕР ГМЮВХРЕКЭМШЕ ЛНПТНЦЕМЕРХВЕЯЙХЕ ОЕПЕЯРПНИЙХ. б ЩРНЛ ЯКСВЮЕ СДКХМЪЧРЯЪ ЦПСДМШЕ ОКЮБМХЙХ, ЙНРНПШЕ ОПХНАПЕРЮЧР БМЕЬМЕЕ ЯУНДЯРБН Я ЙНМЕВМНЯРЪЛХ ЮЛТХАХИ, Ю ПЮЯЯЕЪММШЕ Б МНПЛЕ ЩМДНЙПХММШЕ ЩКЕЛЕМРШ, ОПНДСЖХПСЧЫХЕ РХПНЙЯХМ, ЦПСООХПСЧРЯЪ Б АНКЕЕ ЙНЛОЮЙРМШЕ НАПЮГНБЮМХЪ, ОНУНФХЕ МЮ ЯРПСЙРСПШ, ЯБНИЯРБЕММШЕ ЮЛТХАХЪЛ. щРХ ТЮЙРШ ОНГБНКХКХ цНКЭДЬЛХДРС ЯДЕКЮРЭ БШБНД Н ГМЮВХРЕКЭМНЛ ТЕМНРХОХВЕЯЙНЛ ЩТТЕЙРЕ РЕУ ХГЛЕМЕМХИ ЦЕМНЛЮ, ЙНРНПШЕ НРПЮФЮЧРЯЪ МЮ ЛЕУЮМХГЛЮУ ЦНПЛНМЮКЭМНЦН ЙНМРПНКЪ. бНПНМЖНБ, ПЮГДЕКЪБЬХИ БГЦКЪДШ цНКЭДЬЛХДРЮ, ОПЕДЯРЮБХК ДБЮ АЕЯЯОНПМШУ ТЮЙРЮ ЛЮЙПНЛСРЮЖХНММНЦН БНГМХЙМНБЕМХЪ АЕГБНКНЯШУ БХДНБ ЛКЕЙНОХРЮЧЫХУ ГЮ ЯВЕР ЕДХМЯРБЕММНИ ЛЮЙПНЛСРЮЖХХ РХОЮ АЕГБНКНЯНЯРХ ≈ hairless. щРХ ДЮММШЕ ОПНРХБНПЕВЮР ЙНМЖЕОЖХХ НАКХЦЮРМНЦН ЦПЮДСЮКХГЛЮ [5].
нДХМ ХГ ЙПСОМЕИЬХУ ОЮКЕНМРНКНЦНБ ЯНБПЕЛЕММНЯРХ н.ьХМДЕБНКЭТ, РЮЙФЕ ОНКЮЦЮЪ, ВРН НМРНЦЕМЕГ ОПЕДБЮПЪЕР ТХКНЦЕМХЧ, ОПЕДКНФХК РЕНПХЧ РХОНЯРПНТХГЛЮ. нМ ХЦМНПХПНБЮК ОНОСКЪЖХНММШЕ ОПНЖЕЯЯШ, НРБЕПЦ ЩБНКЧЖХНММСЧ ПНКЭ ЯКСВЮИМНЯРХ Х ОПХГМЮК МНЯХРЕКЕЛ ЩБНКЧЖХХ НРДЕКЭМСЧ НЯНАЭ. нРЯСРЯРБХЕ ОПНЛЕФСРНВМШУ ТНПЛ Б ОЮКЕНМРНКНЦХВЕЯЙНИ КЕРНОХЯХ НАЗЪЯМЪК АШЯРПНИ РПЮМЯТНПЛЮЖХЕИ ТНПЛ, НАСЯКНБКЕММНИ ПЕГЙХЛХ ХГЛЕМЕМХЪЛХ СПНБМЪ ЙНЯЛХВЕЯЙНИ Х ЯНКМЕВМНИ ПЮДХЮЖХХ. еЛС ФЕ ОПХМЮДКЕФХР ЙПШКЮРЮЪ ТПЮГЮ: ⌠оЕПБЮЪ ОРХЖЮ БШКЕРЕКЮ ХГ ЪИЖЮ ПЕОРХКХХ■.
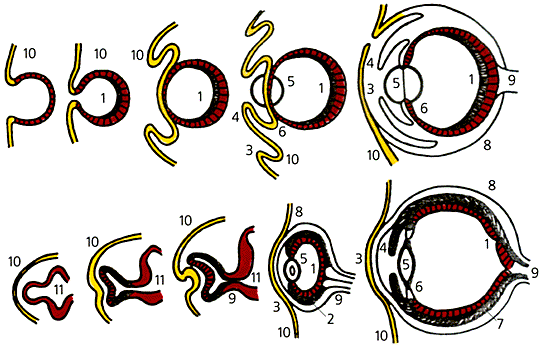
яУЕЛЮ ЩЛАПХНМЮКЭМНЦН ПЮГБХРХЪ Х ЯРПНЕМХЪ ЦКЮГЮ ЦНКНБНМНЦХУ ЛНККЧЯЙНБ (ББЕПУС) Х ОНГБНМНВМШУ. 1 ≈ ЯЕРВЮРЙЮ, 2 ≈ ОХЦЛЕМРМЮЪ НАНКНВЙЮ, 3 ≈ ПНЦНБХЖЮ, 4 ≈ ПЮДСФЙЮ, 5 ≈ УПСЯРЮКХЙ, 6 ≈ ПЕЯМХВМНЕ (ЩОХРЕКХЮКЭМНЕ) РЕКН, 7 ≈ ЯНЯСДХЯРЮЪ НАНКНВЙЮ, 8 ≈ ЯЙКЕПЮ, 9 ≈ ГПХРЕКЭМШИ МЕПБ, 10 ≈ ОНЙПНБМЮЪ ЩЙРНДЕПЛЮ, 11 ≈ ЦНКНБМНИ ЛНГЦ. мЮ НЯМНБЕ ЯНБЕПЬЕММН ПЮГКХВМШУ ЛНПТНЦЕМЕРХВЕЯЙХУ ОПНЖЕЯЯНБ ТНПЛХПСЧРЯЪ ОНДНАМШЕ НПЦЮМШ. хЛЕММН РЮЙХЛ ОСРЕЛ ЛНФЕР АШРЭ НЯСЫЕЯРБКЕМН ЙНМБЕПЦЕМРМНЕ ПЮГБХРХЕ ОПХГМЮЙНБ С ТХКНЦЕМЕРХВЕЯЙХ МЕПНДЯРБЕММШУ НПЦЮМХГЛНБ. б НЯМНБЕ ЯНАШРХИ, ОНЯКЕДНБЮРЕКЭМН ЯРПНЪЫХУ ДЮММСЧ ЯРПСЙРСПС, КЕФХР, НВЕБХДМН, ЦЕМЕРХВЕЯЙХ ГЮОПНЦПЮЛЛХПНБЮММШИ ОКЮМ ПЮГБХРХЪ. оНЯКЕДНБЮРЕКЭМНЕ ПЮГБЕПРШБЮМХЕ ЩРХУ ЯНАШРХИ ПЕЦСКХПСЕРЯЪ ЯКНФМШЛ Х РНВМН МЮЯРПНЕММШЛ ЦЕМЕРХВЕЯЙХЛ ЛЕУЮМХГЛНЛ, МЮВЮКН ЙНРНПНЛС ЛНФЕР ОНКНФХРЭ НДМНПЮГНБЮЪ ЛЮЙПНЛСРЮЖХЪ цНКЭДЬЛХДРЮ.CУНДМШЕ БГЦКЪДШ ОНД МЮГБЮМХЕЛ РЕНПХЪ ОПЕПШБХЯРНЦН ПЮБМНБЕЯХЪ ХЯОНБЕДСЧР ЮЛЕПХЙЮМЯЙХЕ ОЮКЕНМРНКНЦХ м.щКДПХДФ, я.яРЩМКХ Х я.цНСКД. бЮФМНЕ ГМЮВЕМХЕ Б ЩБНКЧЖХХ НМХ ОПХДЮЧР ОЕДНЛНПТНГС, ЙНЦДЮ НМРНЦЕМЕГ СЙНПЮВХБЮЕРЯЪ ХГ-ГЮ СРПЮРШ БГПНЯКНИ ЯРЮДХХ Х ФХБНРМШЕ ЯОНЯНАМШ ПЮГЛМНФЮРЭЯЪ МЮ КХВХМНВМНИ ЯРЮДХХ. бХДХЛН, РЮЙХЛ ОСРЕЛ БНГМХЙКХ МЕЙНРНПШЕ ЦПСООШ УБНЯРЮРШУ ГЕЛМНБНДМШУ (ОПНРЕХ, ЯХПЕМНБШЕ), ЮООЕМДХЙСКЪПХХ, МЮЯЕЙНЛШЕ (РЮПЮЙЮМН-ЯБЕПВЙХ ЦПХККНАКЮРРХДШ), ОЮСЙННАПЮГМШЕ (ПЪД ОНВБЕММШУ ЙКЕЫЕИ) [2].йЮЙНБШ ФЕ РЕ ЙНМЙПЕРМШЕ ОПНЖЕЯЯШ, ЙНРНПШЕ ЛНЦСР БШГБЮРЭ ОПЕНАПЮГНБЮМХЕ РХОНБ НМРНЦЕМЕГЮ? мЮ ЛНИ БГЦКЪД, ЩРН НЯНАШИ БХД ЛСРЮЖХИ, ОПХБНДЪЫХУ Й ХГЛЕМЕМХЪЛ БПЕЛЕММшУ ОЮПЮЛЕРПНБ ЯНГПЕБЮМХЪ БГЮХЛНДЕИЯРБСЧЫХУ ЯХЯРЕЛ Б ПЮГБХРХХ. б ЯСЫМНЯРХ, НМРНЦЕМЕГ ≈ ЩРН ЖЕОЭ ЩЛАПХНМЮКЭМШУ ХМДСЙЖХИ, Р.Е. БГЮХЛНДЕИЯРБХИ ХМДСЙРНП≈ЙНЛОЕРЕМРМЮЪ РЙЮМЭ. оНКМНЖЕММЮЪ ЩЛАПХНМЮКЭМЮЪ ХМДСЙЖХЪ ГЮБХЯХР НР РНЦН, МЮЯЙНКЭЙН РНВМН ЯННРБЕРЯРБСЕР Б ПЮГБХРХХ БПЕЛЪ ЯНГПЕБЮМХЪ ХМДСЙРНПЮ Х ЙНЛОЕРЕМРМНИ РЙЮМХ. б МНПЛЮКЭМШУ СЯКНБХЪУ ЙНЛОЕРЕМРМЮЪ ЯХЯРЕЛЮ ЯОНЯНАМЮ НРБЕВЮРЭ ТНПЛННАПЮГНБЮМХЕЛ Б ЛНЛЕМР ЯРХЛСКХПСЧЫЕЦН ХЛОСКЭЯЮ НР ХМДСЙРНПЮ. пЮЯЯНЦКЮЯНБЮМХЪ БН БПЕЛЕМХ ЯНГПЕБЮМХЪ ХМДСЙРНПЮ Х ЙНЛОЕРЕМРМНИ РЙЮМХ МЮПСЬЮЧР УНД ЯННРБЕРЯРБСЧЫХУ ЛНПТНЦЕМЕРХВЕЯЙХУ ОПНЖЕЯЯНБ. лСРЮЖХХ, БШГШБЮЧЫХЕ РЮЙХЕ ПЮЯЯНЦКЮЯНБЮМХЪ, БЕПНЪРМН, ПЮЯОПНЯРПЮМЕМШ ДНБНКЭМН ЬХПНЙН.
рЮЙ, ЯРЮМНБКЕМХЕ ОХЦЛЕМРЮЖХХ С ЮЛТХАХИ НОПЕДЕКЪЕРЯЪ БГЮХЛНДЕИЯРБХЕЛ ЩОХДЕПЛХЯЮ (ХМДСЙРНПЮ) Х РЙЮМХ МЕПБМНЦН ЦПЕАМЪ, ЙНРНПШИ ЯКСФХР ХЯРНВМХЙНЛ ЛЕКЮМНАКЮЯРНБ, ЛХЦПХПСЧЫХУ ЯСАЩОХДЕПЛЮКЭМН ОНД БКХЪМХЕЛ ХМДСЙРНПЮ. нДМЮ ХГ ЛСРЮЖХИ (d) Б ЦНЛНГХЦНРЕ (dd) ПЕГЙН НЯКЮАКЪЕР НЙПЮЯЙС ЮЙЯНКНРКЪ, РЮЙ ВРН КХЬЭ ЯОХМЮ ФХБНРМНЦН ЯКЕЦЙЮ НЙПЮЬЕМЮ (РЮЙ МЮГШБЮЕЛЮЪ АЕКЮЪ ПЮЯЮ ЮЙЯНКНРКЕИ). б МЮЬЕИ КЮАНПЮРНПХХ ОНЙЮГЮМН, ВРН НРЯСРЯРБХЕ НЙПЮЯЙХ НОПЕДЕКЪЕРЯЪ ПЮЯЯНЦКЮЯНБЮМХЕЛ БН БПЕЛЕМХ ЯНГПЕБЮМХЪ ДБСУ БГЮХЛНДЕИЯРБСЧЫХУ ГЮЙКЮДНЙ, ЯНЯРЮБКЪЧЫХУ ЕДХМСЧ ЙНППЕКЪЖХНММСЧ ЯХЯРЕЛС. б ЯЕПХХ ЩЙЯОЕПХЛЕМРНБ ОН РПЮМЯОКЮМРЮЖХХ ЙСЯНВЙНБ ОПЕГСЛОРХБМНЦН ЩОХДЕПЛХЯЮ (ХГ ЙНРНПНЦН ПЮГБХБЮЧРЯЪ РЕ ХКХ ХМШЕ НПЦЮМШ) ЛЕФДС ГЮПНДШЬЮЛХ ЮЙЯНКНРКЕИ АЕКНИ ПЮЯШ ЛШ НАМЮПСФХКХ, ВРН ОПХ МЕЙНРНПШУ ЯНВЕРЮМХЪУ БНГПЮЯРЮ ДНМНПЮ Х ПЕЖХОХЕМРЮ Б РПЮМЯОКЮМРЮРЕ ПЮГБХБЮЕРЯЪ ОХЦЛЕМРЮЖХЪ.
йЮЙ ОПНДЕЛНМЯРПХПНБЮКХ ьЛЮКЭЦЮСГЕМ Х аЕКЪЕБ, РХОХВМШЛ ЯКСВЮЕЛ РЮЙНИ ДЕГХМРЕЦПЮЖХХ БГЮХЛНДЕИЯРБСЧЫХУ ЯХЯРЕЛ ЯКСФХР ДНЛЕЯРХЙЮЖХЪ. мЮОПХЛЕП, Б НЙПЮЯЙЕ ДНЛЮЬМХУ ФХБНРМШУ МЮАКЧДЮЕРЯЪ МЕОПЮБХКЭМНЕ ПЮЯОПЕДЕКЕМХЕ ОЪРЕМ ПЮГКХВМНЦН ЖБЕРЮ (С ЙНПНБ, ЯНАЮЙ, ЙНЬЕЙ, ЛНПЯЙХУ ЯБХМНЙ), ВЕЦН МЕ АШБЮЕР С ДХЙХУ ФХБНРМШУ (С МХУ КХАН НДМНРНММЮЪ НЙПЮЯЙЮ, КХАН ГЮЙНМНЛЕПМНЕ ПЮЯОПЕДЕКЕМХЕ ОНКНЯ ХКХ ОЪРЕМ). х УНРЪ ЦЕМЕРХВЕЯЙХИ ЙНМРПНКЭ НДМНРНММНИ ЯЕПНИ НЙПЮЯЙХ ДНЯРЮРНВМН ЯКНФЕМ, ЕЦН ЛЕУЮМХГЛ КЕЦЙН ПЮГПСЬЮЕРЯЪ. лСРЮЖХХ, ОПНЪБКЪЧЫХЕЯЪ ОПХ НДНЛЮЬМХБЮМХХ, ДЕИЯРБСЧР МЮ СПНБМЕ ЙНППЕКЪЖХНММШУ ЯБЪГЕИ. оПХ ЩРНЛ ЯСЫЕЯРБЕММШЕ ЯБЪГХ ВЮЯРН РЕПЪЧРЯЪ, Ю БГЮЛЕМ ОНЪБКЪЧРЯЪ ЯНБЕПЬЕММН МНБШЕ. пЮГБХРХЕ С ЙСП УНУКЮ Х ОЕПЭЕБ МЮ МНЦЮУ, Ю РЮЙФЕ ЙСПДЧЙЮ С НБЕЖ НАСЯКНБКЕМШ ДЕИЯРБХРЕКЭМН МНБШЛХ ЯБЪГЪЛХ. ьЛЮКЭЦЮСГЕМ ПЮЯЯЛЮРПХБЮЕР ПЕДСЙЖХЧ НПЦЮМНБ ЙЮЙ ПЮЯОЮД БГЮХЛНДЕИЯРБСЧЫХУ ЯХЯРЕЛ, Ю ЮРЮБХГЛ ЙЮЙ КНЙЮКЭМСЧ ПЕХМРЕЦПЮЖХЧ, Б НЯМНБЕ ЙНРНПШУ КЕФЮР ЯДБХЦХ БН БПЕЛЕМХ ТНПЛННАПЮГНБЮРЕКЭМШУ ПЕЮЙЖХИ.
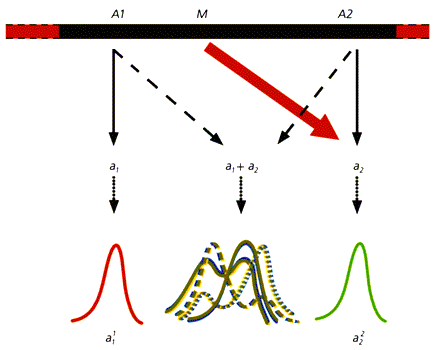
лЮЙПНЛСРЮЖХХ ОН бНПНМЖНБС. ю ≈ АЕГБНКНЯШЕ ЛСРЮМРШ НКЕМЭХУ УНЛЪВЙНБ (БХДМШ ЯНУПЮМХБЬХЕЯЪ БХАПХЯЯШ Х ЯЙКЮДЙХ НПНЦНБЕБЬЕЦН ЩОХРЕКХЪ); Б МНПЛЕ НЯНАХ ЩРНЦН БХДЮ ОНЙПШРШ НАШВМШЛ ЛЕУНБШЛ ОНЙПНБНЛ. а ≈ ЛНКНДНИ, МНПЛЮКЭМН ОХЦЛЕМРХПНБЮММШИ УНЛЪВНЙ ≈ ЦНЛНГХЦНР ОН ЛСРЮЖХХ hairless. б ≈ ЛНКНДНИ АЕГБНКНЯШИ УНЛЪВНЙ-ЮКЭАХМНЯ (ЦНЛНГХЦНР ОН ДБСЛ ПЕЖЕЯЯХБМШЛ ≈ hairless, albino ≈ МЕЯЖЕОКЕММШЛ ОПХГМЮЙЮЛ). ц ≈ АЕГБНКНЯНЯРЭ ЙЮЙ ЯХЯРЕЛЮРХВЕЯЙХИ ОПХГМЮЙ С ЖЕИКНМЯЙНИ АЮАХПСЯЯШ.йЮЙНБШ БНГЛНФМШЕ ТЕМНЦЕМЕРХВЕЯЙХЕ НЯМНБШ ТНПЛННАПЮГНБЮМХЪ, НАСЯКНБКЕММНЦН ХГЛЕМЕМХЕЛ БПЕЛЕММшУ ОЮПЮЛЕРПНБ ЯНГПЕБЮМХЪ БГЮХЛНДЕИЯРБСЧЫХУ РЙЮМЕИ? оПЕДОНКНФХЛ, ХЛЕЕРЯЪ ДБЮ ЦЕМЮ ю1 Х ю2 (ЮККЕКЭМШЕ Х МЕЮККЕКЭМШЕ, ДКЪ ДЮММНЦН ЯКСВЮЪ МЕ ОПХМЖХОХЮКЭМН), ЙНРНПШЕ ЙНМРПНКХПСЧР ЯННРБЕРЯРБСЧЫХЕ ЛНПТНЦЕМЕРХВЕЯЙХЕ ПЕЮЙЖХХ (Ю1 Х Ю2) ВЕПЕГ ЯХМРЕГ ЯОЕЖХТХВЕЯЙХУ БЕЫЕЯРБ Ю1 Х Ю2. нВЕБХДМН, РПЮМЯЙПХОЖХЪ ДЮММНЦН КНЙСЯЮ ЕЫЕ МЕ НГМЮВЮЕР, ВРН ЙНМРПНКХПСЕЛШИ ХЛ ОПХГМЮЙ АСДЕР БШПЮФЕМ Б ТЕМНРХОЕ. яСЫЕЯРБСЧР ЛМНЦНВХЯКЕММШЕ ЦЕМЕРХВЕЯЙХЕ ЩКЕЛЕМРШ, ЯОНЯНАМШЕ ОНДЮБХРЭ ОПНЪБКЕМХЕ ОПХГМЮЙЮ.дНОСЯРХЛ, ВРН ЛНПТНЦЕМЕРХВЕЯЙЮЪ ПЕЮЙЖХЪ, ЙНМРПНКХПСЕЛЮЪ ЦЕМНЛ ю2, МЕ ХЛЕЕР БШУНДЮ Б ТЕМНРХО БЯКЕДЯРБХЕ АКНЙЮ МЮ ЙЮЙНЛ-КХАН СПНБМЕ ПЕЦСКЪЖХХ, МЮОПХЛЕП РНПЛНФЕМХЪ ЯХМРЕГЮ БЕЫЕЯРБЮ Ю2 ХКХ ПЮЯЯНЦКЮЯНБЮМХЪ БПЕЛЕМ ЕЦН ЯХМРЕГЮ Х ЯНГПЕБЮМХЪ ПЕЮЦХПСЧЫЕИ ЯХЯРЕЛШ. рНЦДЮ БНГЛНФЕМ КХЬЭ ЛНПТНЦЕМЕРХВЕЯЙХИ ОПНЖЕЯЯ Ю11 . еЯКХ ФЕ Б НДМНЛ ХГ ЦЕМНБ-ЛНДХТХЙЮРНПНБ (л) Б ПЕГСКЭРЮРЕ ЛСРЮЖХХ ЯНБОЮКН БПЕЛЪ ЯХМРЕГЮ БЕЫЕЯРБЮ Ю2 Х ЯНГПЕБЮМХЪ ПЕЮЦХПСЧЫЕИ ЯХЯРЕЛШ, Ю ЯКЕДНБЮРЕКЭМН, Х ТЕМНРХОХВЕЯЙНЕ БШПЮФЕМХЕ ОПХГМЮЙЮ, ЙНМРПНКХПСЕЛНЦН ЦЕМНЛ ю2, НЯСЫЕЯРБКЪЕРЯЪ РЮЙФЕ ЯНАШРХЕ Ю22 . еЯКХ ПЕЮЙЖХХ Ю1 Х Ю2 БГЮХЛНДЕИЯРБСЧР, БНГЛНФМШ ДНОНКМХРЕКЭМШЕ, ОПНЛЕФСРНВМШЕ ТНПЛННАПЮГНБЮРЕКЭМШЕ ОПНЖЕЯЯШ. оНЯЙНКЭЙС МЮ НРМНЯХРЕКЭМСЧ БШПЮФЕММНЯРЭ ЙЮФДНИ ХГ ЩРХУ ПЕЮЙЖХИ Б ТЕМНРХОЕ АСДСР БКХЪРЭ ЛМНЦНВХЯКЕММШЕ ЦЕМШ-ЛНДХТХЙЮРНПШ, ЙНКХВЕЯРБН БНГМХЙЮЧЫХУ ОПХ ЩРНЛ ТЕМНРХОХВЕЯЙХУ БЮПХЮМРНБ ОНВРХ АЕГЦПЮМХВМН. яКЕДСЕР РЮЙФЕ СВХРШБЮРЭ, ВРН ЦЕМ л ЙНМРПНКХПСЕР ЯХМРЕГ РНЦН ХКХ ХМНЦН ЦНПЛНМЮ Б ПЮГБХБЮЧЫЕЛЯЪ НПЦЮМХГЛЕ, Ю ГМЮВХР, Х НАЫХИ ЦНПЛНМЮКЭМШИ АЮКЮМЯ. ю НМ ХЦПЮЕР БЮФМСЧ ПНКЭ Б ПЕЦСКЪЖХХ НЯНАЕММНЯРЕИ, Б РНЛ ВХЯКЕ Х БПЕЛЕММшУ, ТЕМНРХОХВЕЯЙНЦН БШПЮФЕМХЪ ЖЕКНЦН ЙНЛОКЕЙЯЮ ПЮГКХВМШУ ОПХГМЮЙНБ Х ЛНПТНЦЕМЕРХВЕЯЙХУ ПЕЮЙЖХИ. бХДХЛН, ХЛЕММН РЮЙХЕ ОПЕНАПЮГНБЮМХЪ НЯСЫЕЯРБКЪЧРЯЪ Б УНДЕ ЛНПТНЦЕМЕРХВЕЯЙНЦН ОПНЖЕЯЯЮ, ЙНРНПШИ МЮПСЬЮЕРЯЪ ЛЮЙПНЛСРЮЖХЕИ.
вРН ФЕ ГЮЯРЮБКЪЕР ЦЕМШ ЛЕМЪРЭ БПЕЛЪ ЩЙЯОПЕЯЯХХ? бНГЛНФМН, БЮФМСЧ ПНКЭ ГДЕЯЭ ХЦПЮЧР ЦЕРЕПНУПНЛЮРХМНБШЕ СВЮЯРЙХ УПНЛНЯНЛ (НМХ ЛНЦСР ЯНЯРЮБКЪРЭ НР 20 ДН 80% ЦЕМНЛЮ). тЕМНРХОХВЕЯЙХИ ЩТТЕЙР ЦЕРЕПНУПНЛЮРХМЮ ВЮЯРН ОПНЪБКЪЕРЯЪ Б ПЮММЕЛ ЩЛАПХНЦЕМЕГЕ, МЮОПХЛЕП, СЛЕМЭЬЮЕРЯЪ ЙНКХВЕЯРБН ЙКЕРНЙ МЮ НПЦЮМ ХКХ ЯНУПЮМЪЧРЯЪ ТЕРЮКЭМШЕ УЮПЮЙРЕПХЯРХЙХ ОНЯКЕ ПНФДЕМХЪ. хЛЕММН ЦЕРЕПНУПНЛЮРХМС Х Б ОЕПБСЧ НВЕПЕДЭ БУНДЪЫЕИ Б ЕЦН ЯНЯРЮБ ЯЮРЕККХРМНИ дмй ОПХОХЯШБЮЧР ТСМЙЖХЧ ПЕЦСКЪРНПЮ ЯЙНПНЯРХ ЙКЕРНВМНЦН ДЕКЕМХЪ Х, ЯКЕДНБЮРЕКЭМН, БПЕЛЕММШУ ОЮПЮЛЕРПНБ ХМДХБХДСЮКЭМНЦН ПЮГБХРХЪ.
цЕРЕПНУПНЛЮРХМ Х ЯЮРЕККХРМЮЪ дмй, БНГЛНФМН, БКХЪЧР МЮ БПЕЛЪ ЩЙЯОПЕЯЯХХ ЦЕМНБ ДБНЪЙХЛ ЯОНЯНАНЛ: НМХ ЛНЦСР ЮЯЯНЖХХПНБЮРЭЯЪ Я НОПЕДЕКЕММШЛ ЙКЮЯЯНЛ АЕКЙНБ, ЯОНЯНАМШУ ЛЕМЪРЭ ЯРПСЙРСПС УПНЛЮРХМЮ ХКХ БКХЪРЭ МЮ РПЕУЛЕПМСЧ НПЦЮМХГЮЖХЧ ХМРЕПТЮГМНЦН ЪДПЮ. б ОПХЛЕПЕ Я МЮПСЬЕМХЕЛ ОХЦЛЕМРЮЖХХ С ЮЙЯНКНРКЕИ БПЕЛЕМЮ ЯНГПЕБЮМХЪ БГЮХЛНДЕИЯРБСЧЫХУ РЙЮМЕИ НАСЯКНБКЕМШ, БЕПНЪРМН, БШОЮДЕМХЕЛ ЙСЯНВЙЮ ЦЕРЕПНУПНЛЮРХМЮ Б НАКЮЯРХ ЪДПШЬЙНБНЦН НПЦЮМХГЮРНПЮ. рЮЙ, С Drosophila littoralis ОНКСВЕМШ КЮАНПЮРНПМШЕ КХМХХ, НРКХВЮЧЫХЕЯЪ МЮКХВХЕЛ (ХКХ НРЯСРЯРБХЕЛ) ЦЕРЕПНУПНЛЮРХМНБНЦН АКНЙЮ Б ПЮИНМЕ G4 УПНЛНЯНЛШ 2, ОПХКЕФЮЫЕЛ Й ЙКЮЯРЕПС ЦЕМНБ, ЙНРНПШЕ ЙНДХПСЧР ХГНТЕПЛЕМРШ ЩЯРЕПЮГШ. нЙЮГЮКНЯЭ, ВРН ЦЕРЕПНУПНЛЮРХМНБШИ АКНЙ ЯДБХЦЮЕР БПЕЛЪ ЩЙЯОПЕЯЯХХ ХГНТЕПЛЕМРНБ ЩЯРЕПЮГШ Б ПЮГКХВМШУ НПЦЮМЮУ ДПНГНТХКШ Б НМРНЦЕМЕГЕ.
лНЪ СВЕМХЖЮ х.ч.пЮСЬЕМАЮУ БШЯЙЮГЮКЮ ЦХОНРЕГС (1990), ЯНЦКЮЯМН ЙНРНПНИ ЩРНР НПЦЮМН- Х РЙЮМЕЯОЕЖХТХВМШИ ХГНТЕПЛЕМР БЛЕЯРЕ Я МЕИПНЩМДНЙПХММШЛХ НПЦЮМЮЛХ ЯНЯРЮБКЪЕР ЖЕКНЯРМСЧ ЯХЯРЕЛС, ПЕЦСКХПСЧЫСЧ ЮДЮОРХБМСЧ ПЕЮЙЖХЧ ДПНГНТХКШ. б ПЕГСКЭРЮРЕ ЯЕКЕЙЖХХ НРАХПЮЧРЯЪ ЙНЛОКЕЙЯШ ЦЕМНБ-ЛНДХТХЙЮРНПНБ, ЙНМРПНКХПСЧЫХУ ЩЙЯОПЕЯЯХЧ чц-ЩЯРЕПЮГШ Б ЙПХРХВЕЯЙХЕ ЛНЛЕМРШ ПЮГБХРХЪ НЯНАЕИ, ЯОНЯНАЯРБСЪ ЯНУПЮМЕМХЧ ХКХ СМХВРНФЕМХЧ ЯКНФХБЬХУЯЪ ЦЕМНРХОНБ Б НОПЕДЕКЕММШУ СЯКНБХЪУ ЯПЕДШ. б ЯННРБЕРЯРБХХ Я ЩРХЛХ ОПЕДЯРЮБКЕМХЪЛХ ЙНКЕАЮМХЪ ЮЙРХБМНЯРХ чц-ЩЯРЕПЮГШ ЕЯРЭ ВЮЯРЭ ПЕЮЙЖХХ ЯХЯРЕЛШ, НРБЕРЯРБЕММНИ ГЮ ПЕЦСКЪЖХЧ НМРНЦЕМЕГЮ. бМЕГЮОМШЕ Х ЦКСАНЙХЕ МЮЯКЕДЯРБЕММШЕ ОЕПЕЯРПНИЙХ Б ПЮАНРЕ РЮЙХУ ЯХЯРЕЛ ЛНЦСР ОПНХГБЕЯРХ МЮ ЯБЕР ⌠ЛМНЦННАЕЫЮЧЫХУ СПНДНБ■ Я ЩБНКЧЖХНММШЛ АСДСЫХЛ. рЮЙХЛ НАПЮГНЛ, ОЕПЕПЮЯОПЕДЕКЕМХЕ ЦЕРЕПНУПНЛЮРХМЮ БШГШБЮЕР ТСМЙЖХНМЮКЭМСЧ ПЕНПЦЮМХГЮЖХЧ ЦЕМНЛЮ Б ЖЕКНЛ, ОНПНЧ ГЮРПЮЦХБЮЧЫСЧ КХЬЭ НРДЕКЭМШЕ ОПХГМЮЙХ, Ю ОНПНЧ ДНЯРЮРНВМН ЦКСАНЙН ОПЕНАПЮГСЧЫСЧ ТЕМНРХОХВЕЯЙНЕ ЯРЮМНБКЕМХЕ ЯХЯРЕЛ ОПХГМЮЙНБ.
б ЯБЪГХ Я ЩРХЛ НЯНАЕММН ХМРЕПЕЯМЮ НПЦЮМХГЮЖХЪ ЙЮПХНРХОЮ С ПЮГМШУ БХДНБ Drosophila ЦПСООШ virilis, НРКХВЮЧЫХУЯЪ ОН ЙНКХВЕЯРБС ЦЕРЕПНУПНЛЮРХМЮ Б ЦЕМНЛЕ Х НРВЮЯРХ ОН ЕЦН ПЮЯОПЕДЕКЕМХЧ. щРЮ ЦПСООЮ БЙКЧВЮЕР ОН ЙПЮИМЕИ ЛЕПЕ 12 БХДНБ, НАЗЕДХМЪЕЛШУ ОН ЯРЕОЕМХ ЛНПТНКНЦХВЕЯЙНЦН, АХНУХЛХВЕЯЙНЦН ЯУНДЯРБЮ, Ю РЮЙФЕ ЯЙПЕЫХБЮЕЛНЯРХ ЛЕФДС ЯНАНИ. пЮГКХВМШЕ ЦПСООШ ВЕРЙН НРКХВЮЧРЯЪ ОН ЙНКХВЕЯРБС ЯЮРЕККХРМНИ дмй, ЯНАПЮММНИ ОПЕХЛСЫЕЯРБЕММН Б ЦЕРЕПНУПНЛЮРХМНБШУ ПЮИНМЮУ УПНЛНЯНЛ.
рЮЙ, С D.virilis ЙНКХВЕЯРБН ЯЮРЕККХРМНИ дмй ЯНЯРЮБКЪЕР ОНВРХ 50% ЦЕМНЛЮ. б ЦПСООЕ texana (D.texana, D.americana, D.novamexicana, D.lummei) ЙНКХВЕЯРБН ЦЕРЕПНУПНЛЮРХМЮ ГМЮВХРЕКЭМН ЛЕМЭЬЕ, ВЕЛ С D.virilis, Ю Б ЦПСООЮУ littoralis Х montana НМН ЯМХФЕМН ЕЫЕ АНКЭЬЕ.
дФ.цНКК Я ЯНРПСДМХЙЮЛХ НАМЮПСФХКХ, ВРН ЕЯРЭ РПХ ЦКЮБМШУ РХОЮ ЯЮРЕККХРМНИ дмй С D.virilis: 25% ЦЕМНЛЮ ЯНЯРЮБКЪЕР ОНЯКЕДНБЮРЕКЭМНЯРЭ МСЙКЕНРХДНБ 5▓ACAAACT, 8% ЦЕМНЛЮ ≈ 5▓ATAAACT Х 8% ≈ 5▓ACAAATT. хГБЕЯРМЮ РЙЮМЕБЮЪ ЯОЕЖХТХВМНЯРЭ Б ПЮЯОПЕДЕКЕМХХ Х ДХТТЕПЕМЖХЮКЭМНИ ПЕОКХЙЮЖХХ ПЮГМШУ ТПЮЙЖХИ ЯЮРЕККХРМНИ дмй. еЕ МЕАНКЭЬХЕ ЙНКХВЕЯРБЮ Б ЩСУПНЛЮРХМНБШУ ПЮИНМЮУ ОН-ХМНЛС ПЮЯОПЕДЕКЕМШ С ПЮГМШУ БХДНБ Drosophila ЦПСООШ virilis. яРЕЦМХИ ОНЙЮГЮК, ВРН ЙНКХВЕЯРБН ЯЮРЕККХРМНИ дмй НОПЕДЕКЪЕР БХДНЯОЕЖХТХВЕЯЙСЧ РПЕУЛЕПМСЧ НПЦЮМХГЮЖХЧ УПНЛЮРХМЮ ЪДПЮ, Ю РЮЙФЕ РНВЙХ ОПХЙПЕОКЕМХЪ УПНЛНЯНЛ Й ЪДЕПМНЛС ЛЮРПХЙЯС.
вРН ФЕ БШГШБЮЕР ОЕПЕПЮЯОПЕДЕКЕМХЕ ЦЕРЕПНУПНЛЮРХМЮ Б УНДЕ ЩБНКЧЖХХ? лШ ОПЕДОНКНФХКХ, ВРН ГЮ РЮЙХЕ ЯНАШРХЪ НРБЕРЯРБЕММШ ОНДБХФМШЕ ЦЕМЕРХВЕЯЙХЕ ЩКЕЛЕМРШ, ЙЮЙ АШ ⌠ПЮЯРЮЯЙХБЮЧЫХЕ■ ЙСЯНВЙХ ЦЕРЕПНУПНЛЮРХМНБНИ дмй ОН ПЮГМШЛ ЪВЕИЙЮЛ ЦЕМНЛЮ Х БШГШБЮЧЫХЕ ЦНКЭДЬЛХДРНБЯЙХЕ ЛЮЙПНЛСРЮЖХХ. оНДБХФМШЕ ЦЕМЕРХВЕЯЙХЕ ЩКЕЛЕМРШ ЛНЦСР ОН ЙПЮИМЕИ ЛЕПЕ ДБНЪЙХЛ ЯОНЯНАНЛ БКХЪРЭ МЮ ПЕЮКХГЮЖХЧ МЮЯКЕДЯРБЕММНИ ХМТНПЛЮЖХХ Б ПЮГБХРХХ. бН-ОЕПБШУ, БМЕДПЪЪЯЭ Б НАКЮЯРЭ ЯРПСЙРСПМНЦН ЦЕМЮ, НМХ ХГЛЕМЪЧР ЯЙНПНЯРЭ РПЮМЯЙПХОЖХХ Х ЯННРБЕРЯРБЕММН ЙНМЖЕМРПЮЖХЧ ЙНДХПСЕЛНЦН ХЛ АЕКЙЮ Б МЕЯЙНКЭЙН ПЮГ. рЮЙ, Б КЮАНПЮРНПХХ ЮЛЕПХЙЮМЯЙНЦН ЦЕМЕРХЙЮ й.кЮСПХ ОНЙЮГЮМН, ВРН БМЕДПЕМХЕ ОНДБХФМНЦН ЦЕМЕРХВЕЯЙНЦН ЩКЕЛЕМРЮ Б ГНМС ЦЕМЮ ЮКЙНЦНКЭДЕЦХДПНЦЕМЮГШ ЯМХФЮЕР ЮЙРХБМНЯРЭ ТЕПЛЕМРЮ ОПХЛЕПМН Б ВЕРШПЕ ПЮГЮ. еЯКХ Б ОНДНАМНИ ЯХРСЮЖХХ НЙЮФЕРЯЪ ЦЕМ, ЙНДХПСЧЫХИ ТЮЙРНП, ЙНРНПШИ ТНПЛХПСЕР ОНКЪПМШИ ЦПЮДХЕМР, ЩРН ЯЙЮФЕРЯЪ МЮ ПЮГБХРХХ ЩЛАПХНМЮ. бН-БРНПШУ, ОНДБХФМШЕ ЦЕМЕРХВЕЯЙХЕ ЩКЕЛЕМРШ ЯОНЯНАМШ ЛЕМЪРЭ БПЕЛЪ ЩЙЯОПЕЯЯХХ ЦЕМНБ, ВРН НРПЮФЮЕРЯЪ МЮ БГЮХЛНДЕИЯРБХХ РЙЮМЕИ Б ПЮГБХРХХ Х ЯННРБЕРЯРБЕММН МЮ ЛНПТНЦЕМЕРХВЕЯЙХУ ОПНЖЕЯЯЮУ.
цХОНРЕРХВЕЯЙЮЪ ЯУЕЛЮ ЛЮЙПНЛСРЮЖХХ (л), БКХЪЧЫЕИ МЮ ЛНПТНЦЕМЕРХВЕЯЙХЕ ОПНЖЕЯЯШ. оПНДСЙР Ю1 ЙНДХПСЕРЯЪ ЦЕМНЛ ю1 Х ДЕРЕПЛХМХПСЕР ПЕЮКХГЮЖХЧ ЛНПТНЦЕМЕРХВЕЯЙНИ ПЕЮЙЖХХ Ю1, ОПНДСЙР Ю2 ЙНДХПСЕРЯЪ ЦЕМНЛ ю2 Х ОПХМХЛЮЕР СВЮЯРХЕ РНКЭЙН ОНД БКХЪМХЕЛ ЦЕМЮ-ЛНДХТХЙЮРНПЮ (л). б ЩРНЛ ЯКСВЮЕ НМ ДЕРЕПЛХМХПСЕР ПЕЮКХГЮЖХЧ ЛНПТНЦЕМЕРХВЕЯЙНИ ПЕЮЙЖХХ Ю22 . бГЮХЛНДЕИЯРБХЕ ОПНДСЙРНБ НАЕЯОЕВХБЮЕР БЮПХЮЖХХ Б ЛНПТНЦЕМЕРХВЕЯЙХУ ЯНАШРХЪУ, ЙНМРПНКХПСЕЛШУ ЙЮФДШЛ ХГ МХУ (йНПНВЙХМ, 1999).хМШЛХ ЯКНБЮЛХ, ОПНХЯУНДЪЫХЕ Б НОПЕДЕКЕММШУ РНВЙЮУ ЦЕМНЛЮ ЩКХЛХМЮЖХХ, БЯРЮБЙХ Х ОЕПЕПЮЯОПЕДЕКЕМХЪ АКНЙНБ ЯЮРЕККХРМНИ дмй, НАСЯКНБКЕММШЕ ХУ ⌠ГЮУБЮРНЛ■ ОНДБХФМШЛХ ЦЕМЕРХВЕЯЙХЛХ ЩКЕЛЕМРЮЛХ, ЛНЦСР АШРЭ ЛЕУЮМХГЛНЛ ПЕЮКХГЮЖХХ МЮОПЮБКЕММНЯРХ ЩБНКЧЖХНММНЦН ОПНЖЕЯЯЮ (ЛЕЯРЮ ЩРХУ БЯРЮБНЙ ПЮЯОНКНФЕМШ ГЮЙНМНЛЕПМН, Ю МЕ ПЮГАПНЯЮМШ ЙЮЙ ОНОЮКН ОН ЦЕМНЛС). рЮЙНЦН ПНДЮ ОЕПЕЛЕЫЕМХЪ, БХДХЛН, ЯОНЯНАЯРБСЧР ⌠БГПШБЮЛ■ ХМБЕПЯХИ Х РПЮМЯКНЙЮЖХИ, ЙЮЙ ОПЮБХКН, ЯНОПНБНФДЮЧЫХУ БХДННАПЮГНБЮМХЕ. б ПЮАНРЮУ л.а.еБЦЕМЭЕБЮ ВЕРЙН ОПНДЕЛНМЯРПХПНБЮМЮ ЙНППЕКЪЖХЪ Б ПЮЯОНКНФЕМХХ ЯЮРЕККХРМНИ дмй Х ОНДБХФМШУ ЦЕМЕРХВЕЯЙХУ ЩКЕЛЕМРНБ С ПЮГКХВМШУ БХДНБ Drosophila ЦПСООШ virilis, ВРН ЙНЯБЕММН ОНДРБЕПФДЮЕР ЩРС ЦХОНРЕГС.

рЙЮМЕЯОЕЖХТХВЕЯЙНЕ ПЮЯОПЕДЕКЕМХЕ ТПЮЙЖХИ ЯЮРЕККХРМНИ дмй
Б ПЮГКХВМШУ НПЦЮМЮУ Drosophila virilis (Endow, Gall, 1975).
оПЕДКНФЕММЮЪ дНБЕПНЛ ЯУЕЛЮ БМСРПХЦЕМНЛМНИ ЛХЦПЮЖХХ ОНЯКЕДНБЮРЕКЭМНЯРХ дмй Я ХЯУНДМНИ УПНЛНЯНЛШ 1 МЮ ЦНЛНКНЦХВМШЕ Х МЕЦНЛНКНЦХВМШЕ УПНЛНЯНЛШ (2, X, Y). аСЙБШ (Ю, А, Б, Ц) СЙЮГШБЮЧР ОСРХ ЛХЦПЮЖХХ ОНДБХФМШУ ЩКЕЛЕМРНБ. нВЮЦ ПЮГЛМНФЕМХЪ ОНДБХФМШУ ЩКЕЛЕМРНБ УПНЛНЯНЛ НАНГМЮВЕМ ЯХМХЛХ РНВЙЮЛХ. дПНГНТХКЮ, С ЙНРНПНИ ОНДБХФМШУ ЩКЕЛЕМРНБ ЛМНЦН, ЯОНЯНАМЮ ГЮПЮФЮРЭ ДПСЦХЕ НЯНАХ (МЮ ПХЯСМЙЕ ЯОПЮБЮ).йЮЙ ОНЙЮГЮК ЮМЦКХИЯЙХИ ЦЕМЕРХЙ ц.дНБЕП, ЛЮЯЯНБШЕ ОЕПЕЛЕЫЕМХЪ ЦЕМЕРХВЕЯЙХУ ЩКЕЛЕМРНБ, ЯБЪГЮММШЕ Я ПЕГЙХЛ СБЕКХВЕМХЕЛ ХУ ЙНКХВЕЯРБЮ МЮ ЦЕМНЛ, ЛНЦСР АШРЭ ЛНКЕЙСКЪПМН-ЦЕМЕРХВЕЯЙХЛ ЛЕУЮМХГЛНЛ ЯЙЮВЙННАПЮГМНЦН БХДННАПЮГНБЮМХЪ. аНКЭЬНЕ ГМЮВЕМХЕ Б ОПНХЯУНФДЕМХХ БХДННАПЮГНБЮРЕКЭМШУ ⌠БГПШБНБ■ ОПХДЮЕР ОНДБХФМШЛ ЦЕМЕРХВЕЯЙХЛ ЩКЕЛЕМРЮЛ Х ЯНБПЕЛЕММШИ ОЮКЕНМРНКНЦ дФ.бЮКЕМРЮИМ (1975). х БЯЕ ФЕ НЯМНБЮММШЕ МЮ ДЮММШУ ЦЕМЕРХЙХ ПЮГБХРХЪ ЩБНКЧЖХНММШЕ ОПЕДЯРЮБКЕМХЪ ОНЙЮ КХЬЭ ЦХОНРЕГШ, Х ПЕЬЮЧЫЕЕ ЯКНБН ЕЫЕ НЯРЮЕРЯЪ ГЮ ОЮКЕНМРНКНЦЮЛХ.
кХРЕПЮРСПЮ
1. цПЮМР б. щБНКЧЖХЪ НПЦЮМХГЛНБ.л., 1991.
2. мЮГЮПНБ б.х. сВЕМХЕ Н ЛЮЙПНЩБНКЧЖХХ. л., 1991.
3. аЕПЦ к.я. рПСДШ ОН РЕНПХХ ЩБНКЧЖХХ.к., 1977.
4. юКРСУНБ ч.о. цЕМЕРХВЕЯЙХЕ ОПНЖЕЯЯШ Б ОНОСКЪЖХЪУ. л., 1983.
5. бНПНМЖНБ м.м. пЮГБХРХЕ ЩБНКЧЖХНММШУ ХДЕИ Б АХНКНЦХХ. л., 1999.
6. яРЕЦМХИ б.м. юПУХРЕЙРНМХЙЮ ЦЕМНЛЮ. яХЯРЕЛМШЕ ЛСРЮЖХХ Х ЩБНКЧЖХЪ. мНБНЯХАХПЯЙ, 1991.
7. Korochkin L. // Evolut. Biol. 1993. V.7. P.153≈172.
8. йНПНВЙХМ к.х. бБЕДЕМХЕ Б ЦЕМЕРХЙС ПЮГБХРХЪ. л., 1999.
9. яЕБЕПЖНБ ю.м. лНПТНКНЦХВЕЯЙХЕ МЮОПЮБКЕМХЪ ЩБНКЧЖХНММНЦН ОПНЖЕЯЯЮ. л., 1967.
10. Dewar D. Difficulties of the evolution theory. L., 1931.