№5, 2007 г.
© Узбеков Р.Э., Алиева И.Б.
Центросома - концертмейстер клетки Р.Э. Узбеков, И.Б. Алиева
Рустем Эдуардович Узбеков, д.б.н., с.н.с. группы отдела электронной микроскопии
НИИ физико-химической биологии им.А.Н.Белозерского МГУ им.М.В.Ломоносова.
Ирина Борисовна Алиева, д.б.н., с.н.с. того же отдела.Она из тех, в кого влюбляются с первого взгляда, кому хранят верность всю жизнь и кого не понять до конца. Эта так и не разгаданная за 130 лет исследований тайна скрыта в центросоме - крошечной точке в геометрическом центре клетки, куда радиально сходятся микротрубочки (своеобразные рельсы для внутриклеточного транспорта).
Центросому сравнивают с улыбкой Моны Лизы, называют мерцающей звездочкой, центром цитоплазматической вселенной, клеточным концертмейстером и, наконец, центральной загадкой клеточной биологии. Едва ли найдется в живой клетке другая структура, которую исследователи наделили таким количеством романтических эпитетов; и это не удивительно! Одного взгляда в электронный микроскоп достаточно, чтобы заметить, как сильно центросома выделяется на фоне остальных клеточных структур. Особый интерес любого наблюдателя вызывают главные компоненты этой сложно организованной органеллы - центриоли, по форме напоминающие фрагмент античной колонны.
Однако первые исследователи процесса клеточного деления B.Флемминг, O.Гертвиг и Э.ван Бенеден, почти одновременно описавшие центросому в середине 70-х годов XIX в., увидели лишь темные гранулы в обоих полюсах митотического веретена (рис.1). Да иначе и быть не могло, ведь размер этой органеллы находится на пределе разрешения светового микроскопа. В связи с этим в делящихся клетках сначала были описаны две симметрично располагающиеся структуры, имевшие вид "лучистого сияния", - центросферы. Гранулы в фокусах каждой центросферы первоначально были названы полярными корпускулами.
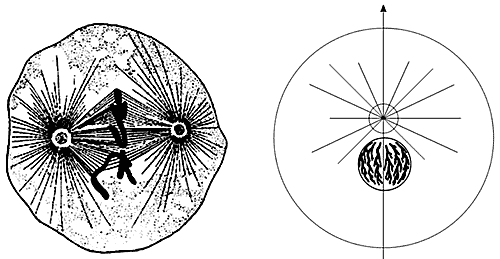
Рис.1. Такой увидели центросому в 1887 г. ее первооткрыватели: Т.Бовери описал ее в полюсах митотического веретена (слева),
а Э.ван Бенеден - в интерфазной клетке.В 1887 г. ван Бенеден вместе с А.Нейтом и независимо от них Т.Бовери установили, что полярные корпускулы полностью не исчезают после деления клетки (митоза). Они сохраняются в течение всего времени между последовательными делениями (этот период жизни клетки теперь называется интерфазой) и при этом часто располагаются вблизи геометрического центра клетки. Ван Бенеден предложил переименовать полярные корпускулы в центральные корпускулы, или центральные тельца, а Бовери - в центросому; он же позднее предложил и термин "центриоль" [1].
Наряду с центросомами, также в конце XIX в., были описаны органеллы, лежащие у основания специализированных клеточных образований - ресничек и жгутиков; эти органеллы получили название кинетосом, или базальных телец [2, 3]. Авторы, Л.Хеннеги и М.Легоссек, наблюдали взаимный переход базальных телец и центросом и в 1898 г. выдвинули гипотезу о гомологии этих клеточных органелл, которая впоследствии получила экспериментальное подтверждение (рис.2).
Рис.2. Формирование полюсов веретена деления из базальных телец в сперматоцитах Bombyx mori [2].
С момента открытия центросомы основное внимание исследователей было приковано к ее роли в организации клеточного деления. После того как Р.Вирхов в 1855 г. сформулировал знаменитый постулат: "Omnis cellula e cellula" ("Каждая клетка от клетки"), исследователи второй половины XIX в. в общих чертах описали картину клеточного деления [4]. Принципиальным для понимания механизма передачи наследственных свойств от клетки к клетке было выяснение роли хромосом. Однако хромосомы сами по себе выглядели пассивными участниками событий митоза, что позволило одному из классиков клеточной биологии, Д.Мезия, сравнить их роль с ролью покойника на похоронах - все происходит ради него, но сам он никакого активного участия в общем действии не принимает. Действительно, при наблюдении митоза в световой микроскоп исследователи видели, как некие нити захватывают хромосомы за их центральные участки и тянут в противоположные стороны клетки. Эти нити были названы нитями веретена (позднее - микротрубочками), а структура, ими образуемая, веретеном деления, поскольку она имела соответствующую форму (рис.2). Оказалось, что нити веретена тянут хромосомы не произвольно, а в направлении строго определенных участков цитоплазмы - полюсов митотического веретена, а в фокусе каждого веретена и располагается главная героиня нашего повествования - центросома!
Хотя центросома с момента ее открытия постоянно находилась в центре внимания биологов, она и более века спустя оставалась, по выражению известного шотландского ученого Д.Уитли, центральной загадкой клеточной биологии [5]. Каким же образом эта едва различимая (занимающая не более 0.1% от общего объема клетки) органелла может выполнять столь важную для жизнедеятельности клетки и организма в целом функцию, как равномерное распределение по дочерним клеткам генетического материала хромосом? Биологи начала ХХ в. предвидели, что центросома, несмотря на малые размеры, устроена не так просто, как кажется на первый взгляд; они надеялись со временем расшифровать ее структуру и тем самым получить ключ к пониманию ее функций. Действительность, как это часто случается, превзошла все, даже самые смелые, предположения первооткрывателей.
Самая обаятельная и привлекательная
Прорыв в исследовании строения центросомы произошел после появления в середине XX в. нового метода исследования - электронной микроскопии. Использование электронного пучка вместо светового луча традиционного микроскопа невероятно расширило возможности морфологического анализа чрезвычайно мелких по величине объектов.
Примечательно, что первое такое исследование центриолей, выполненное С.Селби, оказалось неудачным [6]. Хотя на отдельных микрофотографиях митотических клеток видны косые срезы центриолей, автор не смогла их идентифицировать, а за центриоли приняла осмиофильные гранулы вблизи митотических полюсов. И вот тут весьма кстати оказалась уже упомянутая гомология центриолей и базальных телец, поскольку первые описания ультраструктуры центриолярных цилиндров были сделаны именно на объектах, имеющих жгутики и реснички, - на клетках ресничного эпителия и на сперматозоидах [7, 8]. Сразу после этого была описана и ультраструктура митотических и интерфазных центриолей [9, 10].
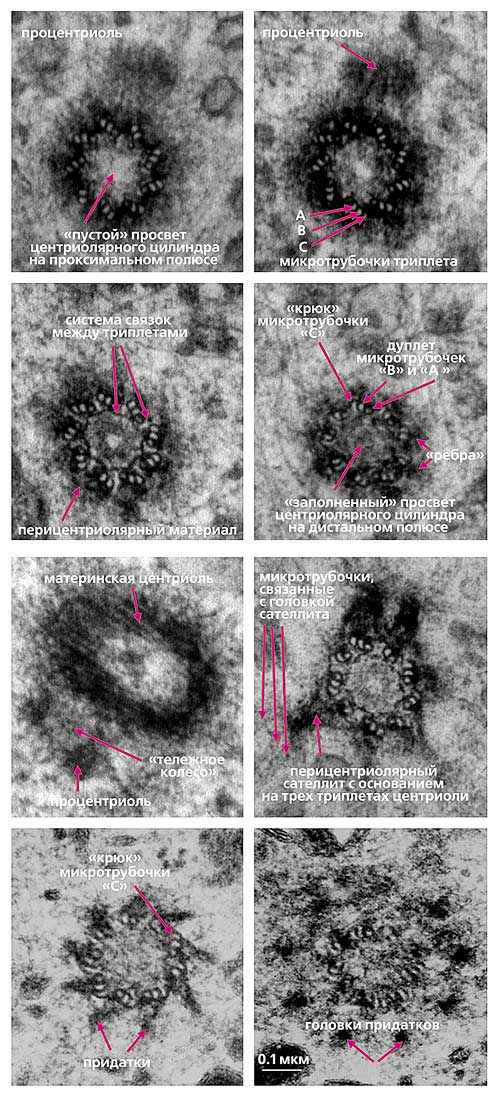
Рис. 3. Ультраструктура центросомы в интерфазной клетке млекопитающих на последовательных серийных срезах [19].Здесь и далее микрофотографии авторов
К настоящему времени ультраструктура центриолей и ассоциированных с ними структур детально исследована. Выяснилось, что в состав центросомы входит пара центриолей, окруженных перицентриолярным материалом (рис.3). Центриоли в паре не одинаковы, одна из них (зрелая, или материнская), в отличие от второй (незрелой, или дочерней), несет на себе дополнительные структуры (рис.3, 4). Оказалось, что созревание центриоли занимает более одного клеточного цикла; в течение первого цикла формирующийся цилиндр, называемый в это время процентриолью, дорастает до нормального размера (см. рис.3, 4).
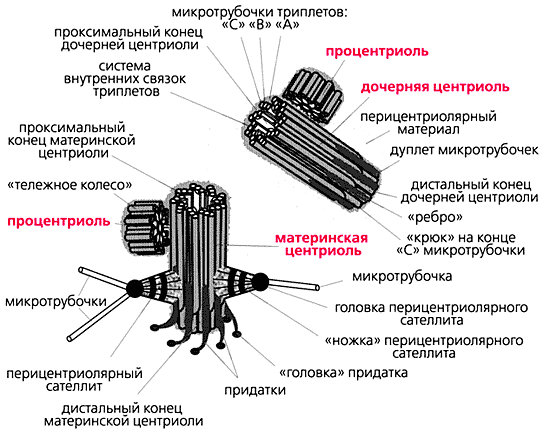
Рис. 4. Упрощенная схема строения центросомы в интерфазных клетках млекопитающих в середине S-фазы клеточного цикла [19].
Длина центриолярных цилиндров составляет 0.3-0.5 мкм, диаметр около 0.2 мкм, при этом стенки их состоят из девяти симметрично расположенных тяжей [7], каждый из которых составлен из трех латерально связанных друг с другом микротрубочек (внутренней - А, средней - B и внешней - С), называемых вместе триплетом [11].
Центриолярный цилиндр - полярная структура. Поскольку в базальном тельце конец цилиндра, от которого растет ресничка, обращен к внешней поверхности клетки, он был назван дистальным, а противоположный конец, обращенный внутрь клетки, - проксимальным. В центриолях придатки и перицентриолярные сателлиты располагаются ближе к дистальному концу, и от него же может расти первичная ресничка (рис.5). В то же время процентриоль (вновь формирующаяся центриоль) всегда образуется ближе к проксимальному концу (см. рис.4). Именно здесь, на проксимальном конце, располагается структура, характерная только для молодых (незрелых) центриолей, - так называемая "ось со спицами", или "тележное колесо" (см. рис.4).
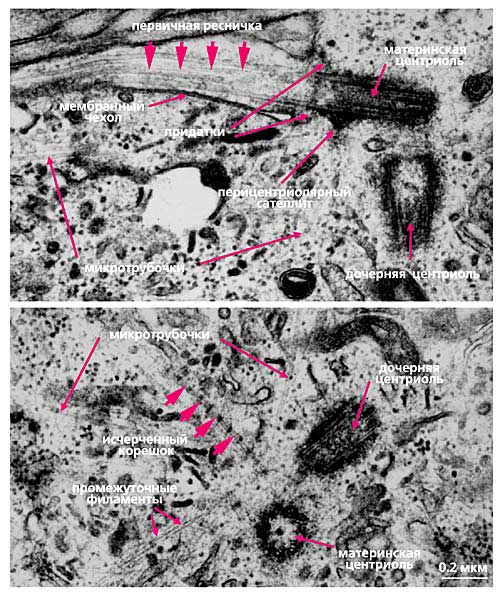
Рис. 5. Ультраструктура первичной реснички исчерченных корешков в интерфазной клетке млекопитающих [20]. © Elsevier Ltd, 2003.
Триплеты микротрубочек лежат под углом к радиусу центриолярного цилиндра, причем закручены они в центриолях всех исследованных объектов одинаково - против часовой стрелки, если смотреть на центриоль с проксимального конца.
Микротрубочки (также полярные биополимеры) в составе центриолярных триплетов всегда ориентированы одинаково - их минус конец находится на проксимальном конце центриолярного цилиндра, а плюс конец - на дистальном.
С поверхностью материнской центриоли связаны структуры двух типов. Во-первых, это перицентриолярные сателлиты (образования, напоминающие по форме фишку детской игры), состоящие из конической ножки длиной около 0.1 мкм, на вершине которой находится округлая головка (см. рис.4). Число их варьирует в норме от одной до четырех на центриоль, но может достигать девяти и более, либо они вовсе отсутствуют в клетках некоторых типов. С головками перицентриолярных сателлитов часто связаны отходящие от центросомы микротрубочки, причем от сателлитов их может отходить значительно больше, чем от стенки центриоли. Перицентриолярные сателлиты - структуры, характерные исключительно для интерфазной центросомы. За несколько часов до митоза они исчезают, и их материал включается в состав так называемого митотического гало - аморфной тонкофибриллярной структуры диаметром около 1 мкм, окружающей центросому в митозе.
Второй тип выростов на поверхности центриолярных цилиндров - придатки, они расположены на дистальном конце каждого триплета, а потому их количество всегда равно девяти (см. рис.4). В отличие от перицентриолярных сателлитов, придатки не исчезают при переходе клетки из интерфазы в митоз, и по их наличию всегда можно определить более зрелую материнскую центриоль.
У материнской центриоли есть еще одна особенность: она способна формировать рудиментарную (первичную) ресничку - структуру, которая выступает над поверхностью клетки подобно реснице над глазом (см. рис.5). Первичные реснички появляются в клетках вскоре после завершения деления и исчезают перед митозом или в самом его начале. С центриолями, формирующими первичную ресничку, часто ассоциированы исчерченные корешки (см. рис.5). Назвали их по предполагаемой функции - первоначально считалось, что они заякоривают ресничку, подобно корням дерева. Однако исчерченные корешки могут наблюдаться и в отсутствие реснички [12].
Строение интерфазной центросомы постепенно меняется в зависимости от стадии клеточного цикла. В конце интерфазы или в профазе митоза две пары центриолей начинают расходиться и формируют два равнозначных центра полимеризации микротрубочек - профазные звезды, при этом интерфазные микротрубочки полностью разрушаются. Каждый полюс веретена в митозе содержит две взаимно перпендикулярных центриоли - диплосому (рис.6). Материнскую центриоль легко отличить от дочерней, поскольку она имеет два свободных конца и окружена митотическим гало.
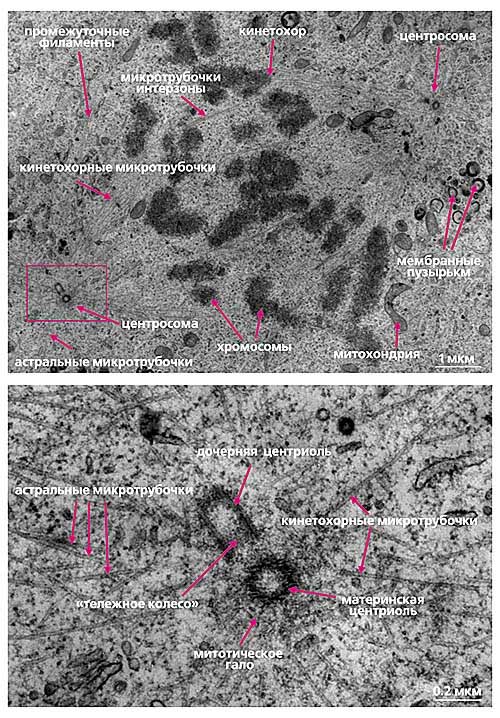
Рис. 6. Ультраструктура центросомы в митотической клетке млекопитающих.
Верхнее фото - общий вид митотического веретена;
нижнее фото - увеличенное изображение диплосомы нижнего левого полюса веретена.
Во всех ты, душечка, нарядах хороша!
По биохимическому составу центросома оказалась мультибелковым комплексом. Первыми, естественно, были охарактеризованы белки, составляющие основу триплетов центриолярных цилиндров, - a- и b-тубулины, а впоследствии семейство пополнилось еще пятью белками - g-, d-, e-, x- и h-тубулинами. Отсутствие любого из них в большей или меньшей степени приводит к нарушению структуры и функций центросомы.
К настоящему времени охарактеризовано уже более сотни ассоциированных с центросомой белков. Поскольку трудно дать единую универсальную классификацию всех этих белков, существует несколько вариантов их систематизации в зависимости от выбранного параметра. По локализации в центросоме различают белки, непосредственно входящие в состав центриолей (как уже упомянутые тубулины), и белки ассоциированных структур и перицентриолярного материала (например, перицентрин). По продолжительности нахождения в центросоме белки разделяют на постоянно присутствующие и появляющиеся в ней только в специфические моменты клеточного цикла. По функциям выделяют несколько групп центросомальных белков: структурные, белки-моторы, регуляторы (в первую очередь киназы и фосфатазы), а также белки, связанные с нуклеацией микротрубочек (образованием затравки, с которой начинается их рост) и удержанием микротрубочек на центросоме.
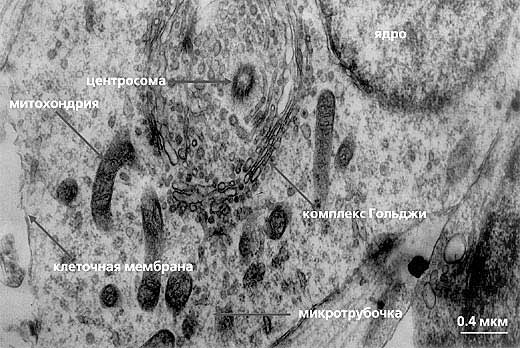
Рис. 7. Центросома, окруженная комплексом Гольджи.
На ультратонком срезе располагается одна центриоль из пары.Белки-моторы, ассоциированные с микротрубочками, участвуют в формировании митотического веретена и осуществляют направленный транспорт вдоль микротрубочек интерфазной сети. При этом микротрубочки выступают в качестве своеобразных рельсов, по которым органеллы и белковые комплексы перемещаются в обоих направлениях - центробежно (от центра клетки к периферии) при участии белков суперсемейства кинезинов, и центрипетально (от периферии клетки к центру) при участии белков суперсемейства динеинов. Необходимо отметить, что центросома часто тесно связана с комплексом Гольджи (рис.7), что обеспечивает доставку созревающих в нем белков по отходящим от центросомы микротрубочкам во все части клетки (рис.8). Регуляторные белки клеточного цикла представлены разнообразными по функциям киназами (осуществляющими специфическое фосфорилирование других белков) - например, киназами CDK1 (p34cdc2), управляющими ходом митоза, или киназами семейств Polo, Aurora, NIMA и др. Белки - компоненты комплекса нуклеации микротрубочек - так же многочисленны, некоторые из них высоко консервативны (т.е. встречаются у всех групп эукариот), другие видоспецифичны. Таким образом, не удивительно, что при столь многообразном белковом составе центросома выполняет в клетке разнообразные функции, часть которых и до настоящего времени полностью не исследована.
На все руки мастерица
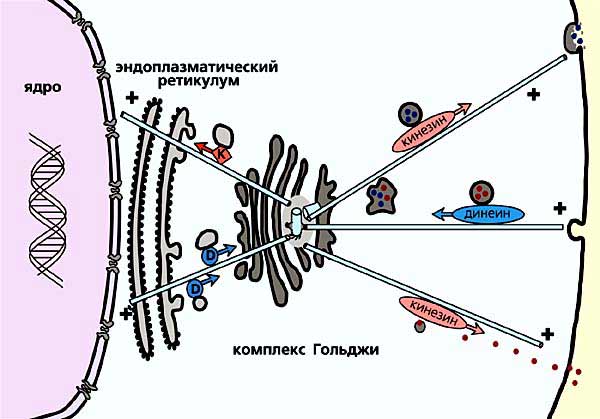
Рис. 8. Схема, иллюстрирующая работу аппарата Гольджи.
Транспорт в направлении к аппарату Гольджи осуществляет моторный белок динеин,
доставку созревших в аппарате Гольджи белков по отходящим от центросомы микротрубочкам
во все части клетки осуществляет моторный белок кинезин.Вспомним, что еще первооткрыватели центросомы связывали ее роль в клетке с функционированием митотического веретена, а значит, и с микротрубочками. Дальнейшие исследования показали, что на центриоли действительно происходит образование (полимеризация) микротрубочек (рис.9), и долгое время полагали, что именно в этом основная функция центросомы. Впоследствии оказалось, что такое представление в значительной степени ограничено, и правы были те исследователи, которые уже в начале XX в. поняли, что эта органелла играет в клетке совершенно особую роль. Однако разберемся с функциями центросомы по порядку.
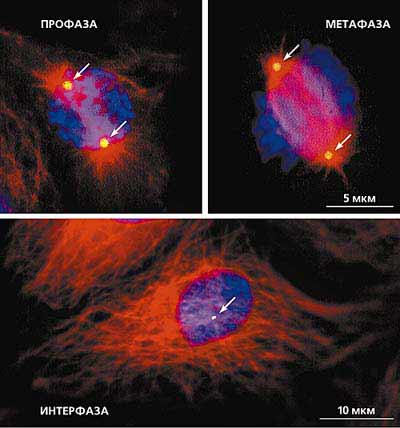
Рис. 9. Центросома и система микротрубочек в профазной, метафазной и интерфазной клетках. Световая микроскопия.
Тройное иммунофлуоресцентное окрашивание выявляет микротрубочки (красный цвет), центросому (зеленый цвет) и ДНК (синий цвет).
Положение центросом показано стрелками.Центросома как центр организации микротрубочек. Это представление о центросоме окончательно оформилось ко второй половине ХХ в. Как было отмечено в обзоре К.Фултон, центросома может организовывать микротрубочки четырьмя различными способами: образует процентриоли, формирует микротрубочки митотического веретена, организует радиальную систему интерфазных микротрубочек, инициирует рост первичной реснички [13]. Созревание центриоли - это и есть не что иное, как приобретение способности к полимеризации микротрубочек [14]. Интересно проследить последовательные стадии, проходя которые центриоль обретает эту способность.
Как мы уже упоминали, окончательное созревание центриоли занимает более одного клеточного цикла. Процентриоли (две на клетку, по одной на каждую уже существующую центриоль) появляются в конце начальной (G1) фазы клеточного цикла и растут на протяжении двух следующих за ней фаз - синтетической (S) и предмитотической (G2). В этом первом для себя клеточном цикле молодые процентриоли не участвуют в нуклеации микротрубочек. Основную роль в формировании их интерфазной системы играет самая старая из четырех центриолей в клетке - "мать" для одной из процентриолей и "бабушка" для другой процентриоли, формирующейся вблизи второй по возрасту центриоли в клетке (см. рис.4).
Далее, в начале митоза, в процессе формирования профазных звезд, центрами нуклеации становятся два митотических гало, в середине которых располагаются диплосомы - структуры, состоящие из ориентированных перпендикулярно друг другу двух центриолей, по одной старой и по одной новообразованной (те самые темные гранулы, наличие которых обнаружили исследователи XIX в.). После окончания митоза дочерняя центриоль оказывается во вновь сформированной клетке в паре с материнской, от которой уже неотличима по размерам. Дочерняя центриоль все еще (в начале G1-фазы второго в своей жизни клеточного цикла) не стала центром организации интерфазных микротрубочек и по-прежнему не может образовывать первичную ресничку (на это тоже способна пока только ее "мать").
Однако в это время молодая дочерняя центриоль впервые отделяется от материнской, и ровно через один цикл после возникновения (в конце G1-фазы второго в своей жизни клеточного цикла) впервые выступает центром организации микротрубочек, формируя новую процентриоль.
В этой связи как нельзя лучше подходит высказанное еще в 1961 г. Д.Мезия предположение: "…когда происходит очередное деление, подготовка к следующему делению уже началась". Более того, можно сказать, что в клетке с закладкой процентриолей началась подготовка не только к ближайшему, но и следующему за ним делению.
При завершении второго клеточного цикла (в профазе митоза) упомянутая центриоль уже может организовывать микротрубочки вторым способом - формировать один из полюсов веретена деления. Тогда же на центриоли появляется белок ценексин. И только прожив в клетке почти два полных цикла, эта центриоль становится, наконец, "старшей" в клетке, центром организации интерфазных микротрубочек и способна формировать первичную ресничку.
Описанный нами сложнейший процесс протекает при участии многочисленных центросомальных белков, многие из которых только ждут своего исследователя. Однако уже понятно, что функции некоторых исследованных белков являются жизненно важными. Так, в начале интерфазы на материнской центриоли формируются перицентриолярные сателлиты. В составе этих органелл обнаружен белок d-тубулин, в отсутствие которого нарушается структура центриолярного цилиндра - происходит потеря микротрубочки "С" и центриоли содержат лишь дуплеты микротрубочек. Без белка центрина невозможно удвоение центриолей. А белок протеинкиназа Аврора А, появляющийся в составе центросомы во второй половине интерфазы, отвечает за регуляцию расхождения центросом (что происходит при участии клеточного белка-мотора Eg5) - будущих полюсов веретена деления.
Мы привели лишь несколько примеров, но и этого достаточно, чтобы понять, насколько значимую роль может играть один-единственный белок в нормальном протекании, тонкой регуляции и филигранно точном исполнении конечного результата столь сложных процессов, в основе которых лежит нуклеация микротрубочек.
Нуклеирующая и заякоривающая функции - две отдельные активности центросомы. Согласно данным последних лет, центросома ответственна не только за нуклеацию микротрубочек, но и за их заякоривание (т.е. закрепление и удержание на центросоме), причем обе функции контролируются разными белковыми комплексами (g-тубулиновым и нинеиновым соответственно) [15]. В клетках культуры ткани оба комплекса расположены в одной локальной области - на центросоме, и это определяет радиальность существующей в них системы микротрубочек. У высокодифференцированных клеток комплексы могут быть сосредоточены в разных участках клетки, что определяет специфическую организацию системы микротрубочек в целом. Например, в эпителиальных клетках, выстилающих орган равновесия (кортиев орган), наряду с расходящимися от центросомы короткими микротрубочками существует множество длинных, ориентированных вдоль длинной оси клетки. Очевидно, что для формирования такой системы микротрубочек необходимо, чтобы заякоривающий комплекс располагался на краю клетки. По-видимому, зародившись на центросоме, короткие микротрубочки перемещаются в направлении клеточной мембраны, откуда дорастают до противоположного конца клетки. Такая специализированная система микротрубочек обеспечивает не только эффективное распределение мембранных компонентов и перемещение везикул, но и выполнение главной специальной функции этих клеток - передачу механических вибраций.
Какие молекулярные механизмы приводят к реорганизации радиальной системы микротрубочек в продольно-ориентированную, до конца неясно. Однако из приведенного примера следует, что радиальная организация сети микротрубочек не универсальна, а центросома не всегда выполняет роль основной структуры, ответственной за пространственную организацию цитоплазматической сети микротрубочек.
Центросома - регуляторный центр клетки. Для этого утверждения есть много оснований, о некоторых из них мы уже говорили, но существуют и другие. Центросома обычно располагается в геометрическом центре клетки, в непосредственной близости от аппарата Гольджи, от нее на периферию клетки радиально расходятся микротрубочки - своеобразные клеточные "рельсы", по которым транспортные молекулы перемещают различные "грузы", а растущая от активной центриоли первичная ресничка выполняет в клетке сенсорную функцию. Считается, что ресничка - элемент пути, транслирующего внеклеточный сигнал на центросому и комплекс Гольджи с целью эффективной секреции новых синтезированных веществ внеклеточного матрикса. Ресничка выполняет роль антенны; на ее поверхности располагаются разнообразные специфические молекулярные комплексы - рецепторы для внешних сигналов. Например, полицистин-2 на поверхности ресничек клеток почечного эпителия участвует в формировании кальциевых каналов и инициации сигнала, контролирующего клеточную пролиферацию и дифференциацию. Одновременно в этих клетках ресничка выполняют и механосенсорную функцию. Рецепторы на мембране реснички могут зависеть от типа клетки - например, реснички нейрона имеют характерные рецепторы для соматостатина и серотонина.
Таким образом, центросома оказывается центральным "узлом" в механизме сигнальной трансдукции: от первичной реснички центросома получает внеклеточный сигнал, в зависимости от которого "регулирует" транспортные процессы, осуществляемые по системе связанных с нею микротрубочек.
Центросома - структурная часть механизма, управляющего динамической морфологией клетки в целом. Живая клетка имеет определенную, характерную для данного типа форму. Форма эта не постоянна, она способна динамично меняться. Постоянство формы клетки поддерживает цитоскелет, и он же обеспечивает ее изменения при различных физиологических и патологических состояниях. Особенно значительные изменения происходят при движении клетки - сложно скоординированном процессе, в который напрямую вовлечены растущие от центросомы микротрубочки. При движении микротрубочки взаимодействуют с актиновым филаментами и клеточными контактами, регулируют натяжение клетки, а изменения их динамики вызывают изменение скорости движения. Выполнение этих функций напрямую связано с пространственной организацией системы микротрубочек, с ее способностью быстро перестраиваться. В настоящее время стала очевидной структурно-функциональная связь всех компонентов цитоскелета в клетке. Так, поддержание формы клетки зависит не только от системы микротрубочек, но и от системы промежуточных филаментов, центр схождения которых также может располагаться вблизи центросомы. Взаимодействие микротрубочек и актиновых микрофиламентов имеет принципиальное значение на различных стадиях построения митотического веретена. Взаимодействие между микротрубочками, актиновыми микрофиламентами и адгезивными структурами является ключевым в регуляции клеточной подвижности (миграции, локомоции, цитокинеза и поляризации клеток). Это взаимодействие осуществляется в первую очередь на структурном уровне посредством белков-связок, которые соединяют микротрубочки и актиновые микрофиламенты [16].
В неспециализированных клетках центросома регулирует не только соотношение свободных и связанных с ней микротрубочек, но и длину радиальных микротрубочек, а, следовательно, и их способность дорасти до края клетки и взаимодействовать своими плюс-концами с фокальными контактами. Дело в том, что единичный растущий конец индивидуальной микротрубочки способен к специфической локальной регуляции контактов путем направленного к ним подрастания микротрубочек - таргетинга [17]. Это делает каждый плюс-конец центросомальной микротрубочки, достигший периферии клетки, потенциально уникальным. Однако способность центросомы сочетать нуклеирующую и заякоривающую функции выходит на первый план не только в связи с представлением о том, что индивидуальная микротрубочка - дискретный инструмент регуляции клеточных контактов, но и в связи с ее способностью закрепляться на специфических сайтах на периферии клетки с помощью комплекса плюс-концевых белков, а также динамически взаимодействовать с актиновыми филаментами [18]. Эта способность плюс-концов очень важна и для митоза, поскольку позволяет радиально растущим от центросомы астральным микротрубочкам взаимодействовать с кортексом и обеспечивать правильное положение ядра, хромосомной пластинки и борозды дробления, а также генерировать силы, действующие на центросому и полюса веретена, с которыми связаны минус-концы микротрубочек. По окончании митоза плюс-концевые белки определяют и положение аппарата Гольджи, в норме локализованного рядом с центросомой; взаимодействие между центросомой и аппаратом Гольджи - необходимый элемент внутриклеточных сигнальных путей регуляции деления клетки и апоптоза.
Мы понимаем, что непосвященному в тайны клеточной биологии трудно воспринять все вышесказанное. Придется поверить на слово: накопленные к настоящему моменту данные свидетельствуют, что центросома - не только центр организации микротрубочек, но и структурная часть механизма, управляющего динамической морфологией клетки в целом.
И вечный бой, покой нам только снится…
Завершая свое краткое повествование о центросоме, попробуем определить, насколько далеко мы продвинулись по пути постижения ее роли в живой клетке. Уникальная центральносимметричная структура всегда вызывала смелые, а порой и фантастические гипотезы о функциях центросомы. История исследований изобилует примерами (большая часть которых, ввиду ограниченности объема, не вошла в данную статью), когда категоричность утверждений исследователей опровергалась сюрпризами, преподносимыми этой клеточной органеллой. Согласно современным представлениям, центросома — важный интегральный элемент живой клетки, функции которой не ограничены ее способностью к полимеризации микротрубочек. В исследовании центросомы появились целые отдельные направления, посвященные ее участию в каком-то одном аспекте жизнедеятельности клетки: в поддержании и изменении формы клетки, в образовании клеточной полярности, в регуляции внутриклеточного транспорта, в формировании мультибелковых ансамблей, ответственных за регуляцию клеточного цикла, и в других клеточных процессах.
Уже на данном этапе развития клеточной биологии понятно, что центросома — ключевая структура в регуляторных процессах, и нарушение ее функций приводит к аномалиям клеточного цикла, дефектам в развитии живых тканей и организмов, к возникновению трофических и онкологических заболеваний. Однако бурное развитие новых экспериментальных подходов дает и, как мы надеемся, будет давать в будущем все новые возможности для исследования центросомы. Несмотря на большое количество описанных центросомальных белков, процесс изучения характера их взаимодействия друг с другом еще только начинается. На наших глазах мозаичность знаний о центросоме сменяется структурированностью, обнаруживаются функциональные связи между различными центросомальными белками. Мощный арсенал молекулярно-биологических и генетических методов в сочетании с детальным изучением морфологии позволяет накапливать огромное количество новых фактов, обработка и анализ которых становятся возможными благодаря современным информационным технологиям. И чем больше мы узнаем о центросоме, тем более важная роль в клетке ей отводится, поэтому без преувеличения можно сказать, что понимание регуляторных функций центросомы как мультибелкового комплекса, видимо, уже в недалеком будущем приведет к более глубокому проникновению в тайны организации живой материи.
Работа выполнена при поддержке Российского фонда фундаментальный исследований.
Проекты №95-04-12703, 00-04-48762, 00-15-99366, 03-04-48035, 06-04-49233.
Литература
1. Boveri T. // Verhandl. Phys. Med. Ges. Wurzburg. 1895. V.XXIX. Цитируется по: [4].
2. Henneguy L.F. // Arch. Microsc. Morph. Exp. 1898. №1. P.481-496.
3. Lenhossйk M.von. Uber Flimmerzellen. Kiel, 1898. V.12. P.106-128.
4. Wilson E.B. The Cell in Development and Inheritance. N.Y., 1900.
5. Wheatley D.N. The Centriole: a central enigma of cell biology. Amsterdam; N.Y., 1982.
6. Selby C.C. // Exp. Cell Res. 1953. V.5. P.386-393.
7. Fawcett D.W., Porter K.R. // J. of Morphology. 1954. №94. P.221-282.
8. Burgos M.H., Fawcett D.W. // J. Biophys. Biochem. Cytology. 1955. V.1. №4. P.287-300.
9. Bernhard W., Harven E.de // Comp. Rend. Acad. Sci. (Paris). 1956. V.242. P.288-290.
10. Yamada T. // Endocr. J. 1956. V.3. №3. P.203-210.
11. Afzelius B. // J. Biophysic. and Biochem. Cytol. 1959. V.5. №2. P.269-278.
12. Alieva I.B., Vorobjev I.A. // Cell Biol. International. 2004. V.28. P.139-150.
13. Fulton C. Centrioles // Origin and continuity of cell organelles. N.Y., 1971. P.170-221.
14. Stubblefield E. // A collection of papers presented the 21 Annual Symposium on Fundamental Cancer Research, 1967. Baltimore, 1968. P.175-193.
15. Mogensen M. Microtubule organizing centers in polarized epithelial cells // Centrosomes in development and disease. Weinheim, 2004. P.299-320.
16. Jaffe A.B., Hall A. // Annu. Rev. Cell. Dev. Biol. 2005. №21. P.247-269.
17. Kaverina I., Krylyshkina O., Small J.V. // J. Cell. Biol. 1999. V.146. №5. P.1033-1044.
18. Bershadsky A.D., Ballestrem C., Carramusa L. et al. // Eur. J. Cell. Biol. 2006. V.85. №3-4. P.165-173.
19. Uzbekov R., Prigent C. // FEBS Letters. 2007. P.1251-1254.
20. Alieva I.B., Vorobjev I.A. // Cell Biology International. 2003. V.28. Р.139-150.