 № 5, 2003 г.
№ 5, 2003 г.
№ 5, 2003 г.
№ 5, 2003 г.
© А.Ю. Журавлев
Невидимые миру факты,
или
«Говорящие» атомы и молекулы в палеонтологииА.Ю. Журавлев
Андрей Юрьевич Журавлев, доктор биологических наук
Нынешняя палеонтология из сугубо описательной науки преображается в систему познания событий, приведших нашу планету в ее теперешнее состояние. Статистические методы переводят бессистемно накопленные материалы в источник полезных сведений [1]. Но это еще не все. Современную палеонтологическую лабораторию трудно представить без множества точных приборов, позволяющих выявить то, что не под силу увидеть даже с помощью мощных микроскопов. Благодаря этим приборам удается разобрать ископаемые раковины и кости на отдельные молекулы и атомы (в основном изотопы).
Конечно, при словосочетании «ископаемая молекула» большинство читателей сразу вспомнит незадачливых героев «Юрского парка», решивших восстановить динозавров по ошметкам ДНК, извлеченным из остатков мезозойского кровососа, влипшего в янтарную смолу. («Клонирование» самого «Парка юрского периода» тоже не очень задалось.) Увы, радужные перспективы извлечения ископаемой ДНК, казавшиеся столь достижимыми всего 10 лет назад, сильно поблекли. Все ДНК миллионолетней давности и даже многие из тех, возраст которых исчисляется тысячелетиями, оказались пресловутыми загрязнениями. Попадание современных молекул (бактериальных, человеческих) в исследуемый материал неизбежно. Эти молекулы, будучи посвежее, и амплифицируются («размножаются») более удачно.
Забавный случай произошел с ДНК из меловых угленосных отложений США. Выделенные из динозавровой кости молекулы отличались от ДНК как млекопитающих, так и пресмыкающихся. «Вот она настоящая ДНК динозавров!» - воскликнули наши американские коллеги, поскорее опубликовали сенсационные данные в ведущих научных журналах и… опять ошиблись. Неведомая ДНК оказалась просто человеческим интроном, как все интроны, мало интересным и поэтому практически неизученным. Теперь представьте себе сюжетец. Молодой ученый с горящими глазами успешно амплифицирует, а затем на лабораторных задворках клонирует такую ДНК, скажем, в геноме лягушки. В результате он получает из пробирки себя любимого или свою лаборантку, тоже любимую, или своего не менее почитаемого учителя. Дальше расписывать можно что угодно: Царевну-лягушку, Пигмалиона, Франкенштейна или Голубую луну на худой конец. (С.Спилберг с М.Крайтоном - создатели книжного и киношного вариантов Юрского парка - отдыхают!)
К сожалению, получить более или менее сносные остатки ДНК из ископаемых материалов возрастом более 100 тыс. лет невозможно. А хорошую (достоверно ископаемую) ДНК редко удается извлечь даже из египетских мумий, которым всего-то немногие тысячи лет. Для палеонтологов - не возраст! Клонировать же подобные остатки и вовсе бессмысленное занятие. Поэтому, когда наши газеты с восторгом пишут, что еще один сибирский мамонт поехал во Францию или Японию для клонирования, - это означает, что он был попросту незаконно вывезен. А существующая исключительно в нашей стране необходимость получить «ветеринарный сертификат» на ископаемые кости - дань (во всех смыслах этого слова) чиновничьему беспределу. Палеонтологический институт РАН не смог, например, ввезти из Монголии ископаемые остатки, которым более 70 млн лет. Не знаю точно, чем руководствовались наши министерства, создавшие сей документ, но это могла быть только самая бульварная фантастика.
И все же бросать поиски хотя бы остатков ДНК не стоит. С помощью ископаемых молекул удалось, скажем, показать, что новозеландский моа не был родственником киви. Это значит, что бескрылые птицы (ратиты) развивались на Новой Зеландии по крайней мере дважды. Сумчатый лев (тилацин) не произошел от южноамериканских сумчатых хищников, а появился независимо от них. Наконец, неандертальца действительно можно считать иным человеческим видом [2].
Однако, кроме ДНК, есть и другие «говорящие» молекулы. С их помощью можно решать не менее интересные задачи. Называются эти молекулы биомаркерами (т.е. следами живого) и давно используются для определения источников и путей миграции нефти. Палеонтологи заинтересовались ими не так давно. Биомаркеры, конечно, представляют собой лишь остатки молекул, но достаточно своеобразные, чтобы восстановить всю цепочку миллионолетних преобразований и вычислить молекулу-предшественницу. Так, хлорофилл а распадается (в разных обстановках) на диоксофиллоэритроэтиопорфирины, этиопорфирины, пристан и фитан. Эти ароматические и алифатические углеводороды могут существовать почти вечно и, будучи извлечены из породы, укажут на присутствие фотосинтезирующих организмов, даже если от них самих не осталось ничего опознаваемого другими методами. Довольно устойчивы остатки жиров. (Проблемы с нерасщепляемыми накоплениями холестерина в виде бляшек всем хорошо известны.)
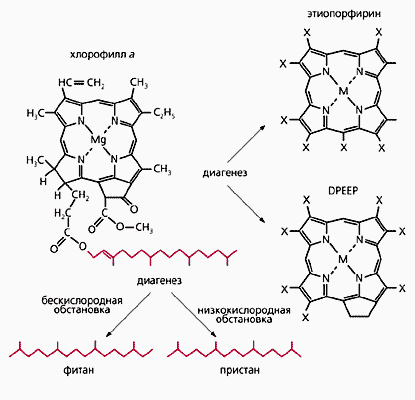
Превращение хлорофилла а в биомаркеры (по М.Молдовану).
DPEEP - диоксофиллоэритроэтиопорфирин, М - ванадий или никель, Х - водород или гидроксильная группа.Только с помощью биомаркеров удалось доказать, что 2.7 млрд лет назад (в архее) на Земле действительно жили не только цианобактерии (о них свидетельствуют метилгопаны), но и эвкариоты [3]. Последних, конечно, выдали остатки жиров - высокоуглеродные стерены, особенно холестен, «привязанный» к холестерину. Древнейшие многоклеточные животные, скорее всего губки, судя по биомаркерам (24-изопропилхолестенам) существовали уже 1.8 млрд лет назад [4]. Это более чем на миллиард лет превышает возраст достоверных губочных скелетных остатков. Сходным образом, хотя и не столь сильно, отличаются по возрасту древнейшие окаменелости цветковых растений (конец раннего мела) и их биомаркеры - олеанины (середина юры). Мало того, такие же биомаркеры извлекли из загадочного пермского растения-гигантоптериса, которое относили к различным голосеменным или их предшественникам [5].
Древние акритархи (слева) и гигантоптерис.
Данный случай показывает, что в настоящее время биомаркеры можно распознавать не только в валовой горной породе, но и в конкретных, даже микроскопических остатках. Так смогли выяснить природу ряда акритарх - загадочных вымерших планктонных организмов. Сначала обнаружилось закономерное совпадение в геологической летописи акритарх, диностеренов и триароматических диностероидов - соединений, характерных для планктонных одноклеточных водорослей- динофлагеллат. А затем эти же биомаркеры получили прямо из акритарховых оболочек [6].
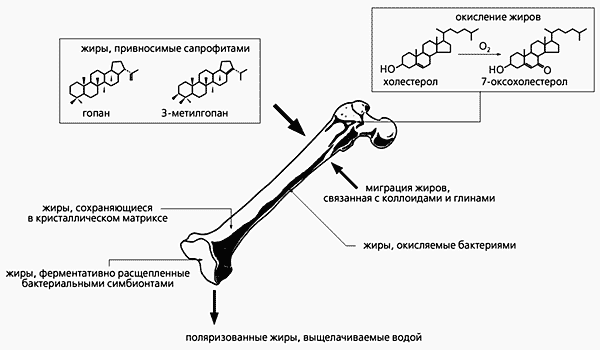
Посмертный цикл жировых накоплений.
Почти все, что мы знаем о земной жизни в течение первых трех миллиардов лет ее развития, установлено благодаря исследованиям биомаркеров и стабильных изотопов углерода, азота, серы и некоторых других элементов. Соотношения устойчивых изотопов в земной коре почти постоянны. Лишь живые существа, сами не отличающиеся особым постоянством, способны нарушить его. Меняется первичное соотношение изотопов, например, за счет фотосинтеза и хемосинтеза. При фотосинтезе из обращения изымается легкий изотоп углерода (12С), который накапливается в органических веществах. Среда, соответственно, обогащается более тяжелым изотопом (13С), что практически навечно запечатлевается в осадке. Обнаружив в горной породе изотопные отличия между органическими остатками (обычно керогенами) и вмещающими отложениями (карбонатами), можно не сомневаться, что к этому «приложили руку» фотосинтезирующие организмы.
Даже если осадок впоследствии будет основательно прокален (при температуре 500°С и давлении в 5000 атмосфер) и первичное содержание изотопов не сохранится, их соотношение изменится закономерным образом. Изотопный сигнал высветил, что жизнь на Земле существует более 3.8 млрд лет [7, 8]. Изотопные микропробы, взятые точно из окаменевших бактерий возрастом 2.1 млрд лет, подтвердили, что это действительно остатки фотосинтезирующих цианобактерий [9]. Иначе как вообще определить, что это за бактерии, если они все на одно «лицо»? (Тут ни сканирующий микроскоп, ни заклинания некоторых наших членов-корреспондентов о пользе «бактериальной палеонтологии» не помогут.)
Изотопная летопись углерода намекает, что содержание в атмосфере метана было высоким вплоть до времени, отстоящего от нашего на 2.7 млрд лет [10, 11]. (Метан, наверное, и служил основным утеплителем Земли, когда Солнце было молодым.) Дело в том, что в органических породах, образовавшихся в морских и наземных (коры выветривания) условиях ранее этого времени, содержится исключительно мало тяжелого изотопа углерода. Значит, метанокисляющие бактерии, наиболее успешно отбирающие легкий изотоп, ни в чем себе тогда не отказывали. Заметный скачок в соотношении изотопов серы (34S/32S) около 2.5 млрд лет назад свидетельствует о переходе ее круговорота под контроль организмов (главным образом сульфатредуцирующих бактерий). А примерно 0.4 млрд лет спустя произошел крупный (на целых 13 единиц) сдвиг в соотношении углеродных изотопов. Его связывают с установлением более кислородной атмосферы и появлением кислородного дыхания, которые не могли не повлиять на окисление органического вещества и, как следствие, на ускоренные темпы его захоронения, что данный сдвиг и отражает. Не случайно в породах этого же возраста начинают встречаться видимые остатки многоклеточных организмов (скорее всего, водорослей). Наконец на границе протерозоя и кембрия (около 550 млн лет назад) изотопные кривые углерода и серы приобрели типичный пилообразный вид, зеркально отражая друг друга. Биогеохимический цикл, вследствие распространения морской макрофауны и особенно организмов, перерабатывающих осадок, приобрел свой прежний вид.
Вообще все крупные рубежи в эволюции биосферы, такие как появление кембрийских скелетных организмов, пермь-триасовое и мел-палеогеновое массовые вымирания, сопровождались резкими скачками в соотношении изотопов серы и углерода, вероятно, вследствие крупных перестроек в трофических цепях.
Изотопы могут поведать и о менее значимых явлениях. Например, о температуре тела динозавров. После находок обширных динозавровых захоронений на Аляске споры о принадлежности динозавров к теплокровным или холоднокровным животным не затухали. Хорошо бы, конечно, измерить температуру у самого динозавра. Оказалось, можно и это, причем без всякого клонирования, а с помощью опять же изотопов [12, 13]. В данном случае - кислорода. Известно, что величины соотношения его стабильных изотопов 18О/16О в осадках (на этот раз не геологических, а метеорологических) закономерно уменьшаются от экваториального пояса к полюсам (промилле на каждые 4°С приповерхностных температур). Происходит так потому, что при испарении водоемы теряют больше легкого изотопа, а в экваториальном поясе этот процесс идет интенсивнее. Животные пьют эту воду из ручьев и луж, и изотопы «застревают» в зубах. Конечно, на самом деле они, пройдя через обмен веществ в организме, откладываются вместе с фосфатом кальция в зубной эмали. В организмах теплокровных (к примеру, млекопитающих) и холоднокровных (скажем, крокодилов) изотопная пропорция приобретает несколько различающиеся величины. В результате прямые линии, проведенные по этим значениям вдоль оси низкие - высокие широты для каждой из названных групп, будут иметь разный наклон. Так вот, прямая, построенная с использованием данных по кислородным изотопам из динозавровых костей, столь же круто наклонена, как у млекопитающих. Для крокодилов же - более полого.
Можно подойти к решению данного вопроса и с другого конца. У теплокровных животных температура примерно одна и та же по всему телу. (Градусник, не боясь ошибиться, можно вставлять и под мышку, и в рот, и еще кое-куда.) Иначе у холоднокровных. У них конечности и другие выступающие части, как правило, холоднее. (Может быть, именно поэтому у китайских пернатых динозавров оперение в первую очередь появилось на лапах и хвосте?) Пускаем в ход уже знакомые изотопы кислорода и по их соотношению меряем температуру в конечностях (фалангах пальцев) и теле (основном скелете) позднемеловых динозавров. Все величины немного отличаются. Юный растительноядный гипсилофодонт больше полагался на внешнее тепло, но перепад температур в частях его тела был меньше, чем у холоднокровных той же размерности. Небольшой протоцератопс оказался довольно «теплокровным», а подросток цератопс сохранял почти постоянную температуру. Крупный (7 - 9 м длиной и около 5 т массой) гадрозавр отличался среди них самым высоким уровнем обмена веществ, т.е. был самым теплокровным. Значит, скорость обмена веществ у позднемеловых динозавров была выше, чем у современных пресмыкающихся, но еще не такой высокой, как у млекопитающих и птиц. Этого как раз достаточно для сохранения почти постоянной температуры тела при внешних изменениях температур в пределах 10 - 15°С, как в безморозном позднемеловом климате.
Соотношение стабильных изотопов углерода в геологической летописи (слева) и степень их фракционирования различными группами организмов.Собственно климат, главным образом температура, тоже восстанавливается по кислородным изотопам. Особенно для тех периодов, когда наземная растительность - самый лучший индикатор палеотемператур - еще не распространилась широко. Хотя живые организмы прямого отношения к разделению этих изотопов не имеют, раковины некоторых видов служат довольно надежным хранилищем первичной изотопной летописи. Так, используя раковины замковых брахиопод, образовавшиеся из низкомагнезиального кальцита, мало подверженного вторичным изменениям, построили кривую падения температур для ордовикского периода. Оказалось, что позднеордовикское оледенение было действительно кратковременным (всего каких-то 500 тыс. лет) и внезапным. Резкое охлаждение планеты, вероятно, и послужило причиной одного из самых массовых вымираний в истории Земли. Выжили в основном холодноводные сообщества, сдвинувшись к экватору.
Самые низкие величины d13С соответствуют степени фракционирования изотопов органического углерода метанобразующими бактериями [7]. Пики на верхней кривой показывают, насколько ускорялись темпы его захоронения в виде карбонатов (когда в атмосфере уже появился кислород) в связи с расколом суперконтинентов и образованием прогибов. 1 - С3-растения, 2 - С4-растения, 3 - метанокисляющие бактерии, 4 - водоросли, 5 - бактерии (а - в естественных местообитаниях, b - в культурах), 6 - анаэробные фотосинтезирующие бактерии, 7 - метанобразующие бактерии (по М.Шидловски). d13С - величины 13С/12С, рассчитанные по отношению к стандарту, которым служит ростр белемнита из меловой формации Пи-Ди (США), Сорг - органические породы, Скарб - карбонаты.Падение температур наблюдается и в конце неогенового периода. Для этого интервала лучшими индикаторами служат раковины фораминифер. Сопоставляя изотопные данные по бентосным и планктонным фораминиферам для разных океанов, можно довольно точно выявить изменения температур и соотнести их с понижением содержания углекислого газа в атмосфере. Этот процесс привел к распространению на суше трав, привычных к дефициту углекислого газа, поскольку они используют несколько иную цепь фотосинтетических реакций. Их называют C4-растениями, так как у них первичные продукты фотосинтеза - четырехуглеродные соединения. К таким растениям принадлежат многие травы тропических и умеренных широт. (Большинство деревьев, кустарников и высокогорных трав относятся к C3-растениям.)
Травы сохраняются в ископаемом состоянии плохо, и о развитии самой продуктивной наземной экосистемы (травянистых ландшафтов) можно судить по изотопам углерода и азота из костей и зубов млекопитающих. Соотношение изотопов углерода отражает, каких растений было больше (C4 или C3), а изотопы азота (15N/14N) дают некоторые представления о продуктивности палеосообщества. Попутно можно отследить пищевые предпочтения давно вымерших растительноядных животных. У тех млекопитающих, которые любят ощипывать древесные листочки (например, у жираф и слонов), в зубах накапливается относительно мало тяжелого изотопа кислорода [14 - 16]. Зато у тех, кто пасется, не поднимая головы (зебры и антилопы-гну и др.), его присутствие заметно.
Если бы не изотопы, мы не только не узнали бы, когда на Земле появились первые фотосинтезирующие организмы, но и когда зазеленела травка. Характерные для трав изотопные соотношения наблюдаются лишь в зубах средненеогеновых млекопитающих. В Южной Америке травянистые ландшафты, возможно, стали обычны даже раньше. (Поскольку все в конечном счете оказывается в море, изменения в соотношении углеродных изотопов проявляются даже в раковинках фораминифер того же возраста.) Однако зубной изотопный расклад не единственное средство поиска того, от чего не сохранилось никаких остатков. Летопись ископаемых почв тоже прекрасно отражает развитие травянистых сообществ. Никакую траву совершенно невозможно оторвать от ее почвы. Изотопные особенности почв свидетельствуют, что в конце палеогенового периода стали появляться пустоши с разбросанными там и сям кустарниками и куртинами трав. (К этому могли приложить тяжелую ногу некоторые растительноядные динозавры - гадрозавры и завроподы.) В начале неогенового периода зацвели маргаритки и бобовые, а к середине раскинулся ковер разнотравья. Сходная цепочка событий выстраивается по углеродным изотопам, извлеченным из растительных биомаркеров.
Вообще, сопоставляя изотопные (атомные) и биомаркерные (молекулярные) данные, можно получить очень интересные результаты. В отложениях Экваториальной Атлантики, накопившихся за последние 200 тыс. лет, проследили, как менялось содержание скелетных остатков кокколитофорид и динофлагеллят, их же биомаркеров (алкенонов и диностеренов, соответственно) и соотношения кислородных изотопов в раковинах планктонных фораминифер. Последние, как упомянуто, показывают изменение палеотемператур. Оказалось, что температурные минимумы очень точно совпадают с максимумами обилия планктонных водорослей [17, 18]. Их биомаркеры не только независимые показатели наличия водорослей, но и степени их продуктивности, которая связана с выделением диметилсульфида. Это вещество, содержащее серу, поступая в атмосферу, распадается на компоненты, способствующие усилению облачного покрова, в результате чего повышается земное альбедо. А это ведет к понижению температуры до уровня, оптимального для роста данных водорослей.
Неужели лавлоковская гипотеза Геи, над которой в последнее время принято смеяться, все-таки не просто гипотеза, и растения (особенно водоросли) - главные регуляторы климата? А почему бы нет? Крупнейшим позднепротерозойским оледенениям предшествовало бурное развитие акритарх (водорослей, содержащих диностерол). Необычное позднеордовикское оледенение, случившееся в условиях «теплой» Земли, сопровождалось всесветным образованием отложений с исключительно обильными остатками водорослей. (Правильнее следует сказать, конечно, наоборот: накопление таких отложений сопровождалось оледенением.) Что каменноугольно-раннепермское похолодание и распространение особого типа лесов совпадали неслучайно, почти никто уже не оспаривает. Нужно только придумать, как получить независимые данные, подтверждающие или опровергающие предположение о тесной связи растительности с климатом и о ведущем факторе в этой связи.
Может быть, опять помогут биомаркеры и изотопы? Пока же они подсказывают, что эвтрофные условия вполне могли привести к позднедевонскому массовому вымиранию. Что доломит, прежде считавшийся исключительно диагенетическим минералом, образуется также биогенным путем в естественных условиях. Что прямоходящие двуногие приматы проживали в высокогорном лесу прежде, чем выйти на степные просторы. И много чего еще. Было бы желание узнать.
Литература
1. Журавлев А.Ю. // Природа. 2001. №9. С.51 - 56.
2. Wayne R.K., Leonard J.A., Cooper A. // Annu. Rev. Ecol. Syst. 1999. V.30. P.457 - 477.
3. Brocks J.J. et al. // Science. 1999. V.285. P.1033 - 1036.
4. McCaffrey M.A. et al. // Geochim. Cosmochim. Acta. 1994. V.58. P.529 - 532.
5. Молдован М. Доклад на XXXI сессии. Международного геологического конгресса. Бразилия, 2000.
6. Moldowan J. M. et al. Molecular fossils demonstrate Precambrian origin of dinoflagellates // The ecology of the Cambrian radiation / A.Yu.Zhuravlev, R.Riding. N.Y., 2000. P.474 - 493.
7. Schidlowski M. et al. // Geochim. Cosmochim. Acta. 1979. V.43. P.189 - 199.
8. Mojzsis S.J. et al. // Nature. 1996. V.384. №6604. Р.55 - 59.
9. House C.H. et al. // Geology. 2000. V.28. №8. P.707 - 710.
10. Hayes J.M. Global methanotrophy at the Archean-Proterozoic transition // Early life on Earth / S.Bengtson. N.Y., 1994. P.220 - 236.
11. Pavlov A.A. et al. // Geology. 2001. V.29. №11. P.1003 - 1006.
12. Barrick R.E., Showers W.J. // Palaios. 1996. V.11. №2. P.295 - 305.
13. Frieke H.C., Rodgers R.R. // Geology. 2000. V.28. №9. P.799 - 802.
14. Cormie A.B., Schwarcz H.P. // Palaeogeogr., Palaeoclimat., Palaeoecol. 1994. V.107. P.227 - 241.
15. Retallack G.J. // Palaios. 1997. V.12. №4. P.380 - 391.
16. MacFadden B.J. // Annu. Rev. Ecol. Syst. 2000. V.31. P.33 - 59.
17. Huang Y. et al. // Science. 2001. V.293. №5535. P.1647 - 1651.
18. Henriksson A.S. et al. // Geology. 2000. V.28. №6. P.499 - 502.
Апрель 2003

