№ 5, 2003 г.
№ 5, 2003 г.
№ 5, 2003 г.
№ 5, 2003 г.
© Ю.Т. Дьяков
ГРИБЫ И РАСТЕНИЯ Ю.Т. Дьяков
Юрий Таричанович Дьяков, доктор биологических наук,
зав. каф. микологии и альгологии биофака МГУ им.М.В.Ломоносова.Важнейший критерий, позволивший Р.Х.Уиттейкеру выделить грибы в отдельное царство биоты, — это осмотрофное питание, причем в большинстве случаев (и для паразитов, симбионтов и сапротрофов) растительными тканями. Ферментативный аппарат грибов (гидролитические ферменты) нацелен на разложение углеводов — строительного материала и запасных веществ растений. Связи грибов с растениями, во-первых, очень древние (видимо, симбиоз этих организмов и обеспечил их выход на сушу) и, во-вторых, весьма разнообразные. Их можно свести к двум главным типам: взаимоотношениям грибов с живыми растениями и деструкциям мертвых растений.
Взаимодействие грибов с живыми растениями
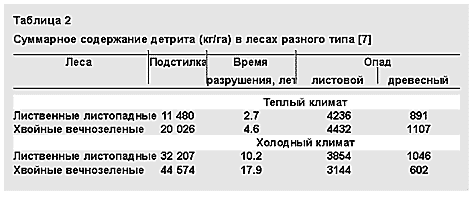
Исторически самые древние ассоциации грибов с растениями — микоризы. Первая из них, найденная в отложениях древнейших наземных растений ринофитов, — арбускулярная микориза. При ней грибной мицелий располагается в межклеточном пространстве корня и образует в его клетках трофические органы — сильно разветвленные выросты, арбускулы. Отсюда и название этой микоризы.
Что же получили растения от такого содружества? Прежде всего улучшилось корневое питание, поскольку увеличился объем почвы, охватываемой всасывающей поверхностью. Радиус грибной гифы на два порядка меньше радиуса корневого волоска, поэтому поверхность корня на единицу объема (или веса) в 100 раз меньше поверхности гифы, т.е. для продукции одинаковой всасывающей зоны растению требуется в 100 раз больше материала, чем грибу.
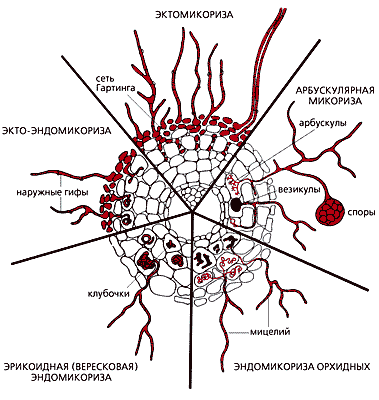
Морфологические особенности различных типов микоризы.
В центре — поперечный срез корня.При таком симбиозе растения получили дополнительное питание в виде элементов, находящихся в труднодоступной форме (фосфаты). Возникла способность к межклеточной регуляции развития микроорганизмов в кортексе корня, к образованию внутриклеточных симбиотических структур, а также к усвоению хитиноподобных метаболитов микросимбионта.
Без микоризы не было бы и симбиоза с клубеньковыми бактериями. Оба процесса — образование клубеньков и грибных арбускул — контролируются перекрывающимися генами растения. Недавно показано, что гены бобовых, продукты которых обеспечивают симбиотическую рецепцию SYMRK (symbiosis receptor-like kinase), контролируют белки со структурой, характерной для R-белков растений, выполняющих иммунные функции [1]. Отсюда — возникновение механизмов химической защиты растений, ограничивших «притязания» симбионтов. В частности, появились такие производные фенолов, как флавоноиды, не только противостоящие патогенам, но и расширившие адаптационные способности растений.
Следующий тип эндотрофной микоризы — эрикоидная — симбиоз сумчатых грибов с вересковыми растениями, позволивший им освоить бедные почвы в экстремальных условиях (в тундрах, горах и др.).
Значительно позднее, в мезозое, возникла эктотрофная микориза, вероятно, образованная базидиальными древоразрушающими грибами. Некоторые виды и сейчас формируют микоризу в ненарушенных лесных ценозах, а при антропогенных воздействиях они вызывают корневые и стволовые гнили. Эволюция эктотрофной микоризы связана с почвообразованием. Как защита от нападения базидиальных грибов появился полимер ароматических веществ — лигнин, который стал играть первостепенную роль в глобальном цикле углерода. Его масса — 1011 т углерода, что составляет 25% от общей фотосинтетической продукции. В лигнине содержится больше углерода, чем в целлюлозе и других полисахаридах, значит, он основной участник и в образовании торфа, почвенного гумуса и, естественно, древесного угля. Эктотрофная микориза обеспечила еще одну важную для фитоценоза возможность: коммуникацию между его отдельными членами и связь между корневыми системами через грибной мицелий. В опытах с использованием радиоактивно меченных элементов питания доказан взаимный обмен метаболитов [2]. Поскольку эктомикоризные грибы слабо специализированы и заражают корни многих видов растений, лесной биоценоз благодаря связям через микоризу можно сравнить с единым организмом.

Эктомикоризная сеть, обеспечивающая контакты между растущими поблизости сеянцами сосны.
Грибы — паразиты растений
Параллельно возникли и другие ассоциации с растениями биотрофных и некротрофных экзо- и эндофитных паразитов. В природе вследствие длительной коэволюции они — источники эндемичных болезней, которые всегда присутствуют в растительных популяциях. Однако их вред невелик, так как, во-первых, у хозяев возникает к ним устойчивость, а во-вторых, накапливаются факторы, снижающие вирулентность самих паразитов. Первая причина скорее всего характерна для однолетних, быстро эволюционирующих растений, а для популяций медленно эволюционирущих многолетних древесных растений — вторая. Например, вирулентность возбудителя рака каштанов Cryphonectria parasitica уменьшается благодаря миграции в популяциях паразита самореплицирующейся двухцепочной РНК. В присутствии такого «гиповируса» не функционирует один из двух генов a-субъединицы G-белка (гетеромерный белок, связывающий гуанозинтрифосфат), прекращаются индукция гена целлобиогидролазы (cbh1) и секреция фермента целлюлазы, необходимого для заражения. Полагают, что фосфорилированный a-белок активирует аденилциклазу и влияет тем самым на передачу сигнала к экспрессии генов (в частности cbh1), участвующих в патогенезе.
Зона эндемичных болезней в отдельные периоды втягивается и выпячивается, но иногда, при очень благоприятных погодных условиях, они могут перерасти и в эпидемию. Среди многих изученных механизмов толерантности растений к эндемичным болезням важную роль играет высокий полиморфизм. Он обеспечивает существование в популяции особей с разной устойчивостью. Когда ботаники впервые применили биохимические методы популяционного анализа растений, в частности анализ изоферментов, они были поражены, сколь высок скрытый генетический полиморфизм даже у самоопыляемых видов, которые, казалось бы, должны эволюционировать в направлении гомозиготности. Важнейшей причиной сохранения высокого генетического полиморфизма у самоопылителей не без основания считают сосуществование с грибными паразитами.
Эндофиты
Конечный результат ассоциации растений и паразитов — сумчатые и базидиальные эндофиты. В их взаимодействиях с растениями наблюдаются все переходы от паразитизма к мутуализму, при котором отношения между партнерами взаимовыгодны и раздельное существование невозможно.
Во-первых, многие эндофиты повышают жизнеспособность зараженных растений: усиливается фотосинтез и продуктивность; ускоряются прорастание семян и рост, повышается устойчивость к стрессам.
Во-вторых, изменяется онтогенез хозяев. В частности, многие сумчатые и большинство базидиальных эндофитов резко снижают семенную продукцию растений; а при неполной стерилизации — заменяют перекрестное опыление на самоопыление. Они активизируют рост, кущение и вегетативное размножение. Например, осоки из рода Cyperus, зараженные сумчатыми грибами из рода Balania, часто не образуют семян, но дают много клубеньков и луковичек. Значит, гипотезы об эндофитном происхождении запасных органов растений (клубеньков и луковиц), высказанные некоторыми ботаниками еще 100 лет назад, не столь уж беспочвенны. Заменяя перекрестное опыление самоопылением, а половое размножение вегетативным, грибы сужают популяционное разнообразие своих хозяев, постепенно вытесняя устойчивые генотипы, заменяя их уже зараженными, т.е. восприимчивыми.
И наконец, сумчатые эндофиты злаков, относящиеся к спорыньевым, образуют в своих хозяевах алкалоиды. Эти вещества придают растениям горький вкус и оберегают их от травоядных животных (от насекомых и слизней до жвачных). Такая плата, которую гриб вносит за пансион с питанием, создает серьезные проблемы в пастбищном скотоводстве, ибо зараженные травы вызывают заболевания скота. Вместе с тем эндофиты, способствующие интенсивному кущению, защите от вредителей и болезней, весьма перспективны в селекции и разведении газонных трав.
Роль грибов в стабилизации фитоценозов
Известный зоолог Н.Н.Беклемишев считал паразитов важным стабилизирующим компонентом биоценозов [3]. Рассмотрим, как влияют грибы на гомеостаз фитоценозов.
Прежде всего они формируют биологическое разнообразие. Поскольку степень пораженности зависит от густоты стояния растений, наиболее чувствительны к патогенам конкурентоспособные и, следовательно, плотно растущие растения. Снижение их жизнеспособности (продуктивности фотосинтеза, урожая и всхожести семян) под влиянием болезни позволяет менее конкурентоспособным видам сосуществовать с ними в популяции. При формировании сложных фитоценозов фитопатогенные грибы выступают как регуляторы численности его сочленов. Например, гриб Stachybotrys chartarum вызывает корневую гниль проростков ветренницы дубравной Anemona nemorosa. В результате примерно 20% семенных проростков погибает, не достигая поверхности почвы. У звездчатки Ustilago violacea возбудитель головни, стерилизуя цветки, резко снижает семенную продукцию зараженной популяции. Ржавчинный гриб Gymnosporangium juniperinum паразитирует на можжевельнике и рябине. Развиваясь в лесах, верхний ярус которых образует рябина, а подлесок — можжевельник, ржавчина вызывает частичную дефолиацию рябины и просветление леса, обеспечивающее рост можжевельника. Разнообразию фитоценозов способствуют не только паразитические, но и микоризные грибы.
Недавно было высказано соображение (принцип Янцена—Коннела) о роли патогенов в пространственном распределении растений в фитоценозах, которое уже экспериментально подтвердилось. Например, в лесах Канады всходы дикой вишни около породившего их дерева поражаются сильнее, чем вдали от него [4]. Объяснение простое: патогенные штаммы рода Pythium, вызывающие корневые гнили, накапливаются вблизи деревьев, причем степень их влияния тем сильнее, чем больше густота всходов.
Грибы обеспечивают сукцессию видов растений в бедных фитоценозах. На песчаных дюнах вдоль побережья Северо-Западной Европы растут многолетние злаки и осоки, размножающиеся клонально (видимо, вследствие присутствия эндофитных грибов). Развитие каждого вида проходит через несколько этапов: колонизацию, оптимальный рост, дегенерацию, замену другим видом. Причины такой сукцессии — разная степень толерантности к погодным условиям и к возбудителям корневых гнилей. При замене каждый новый вид чувствителен к заражению микроорганизмами, выделенными из почвы в области его корневой системы, но устойчив к микроорганизмам из зоны корневой системы предыдущего вида.
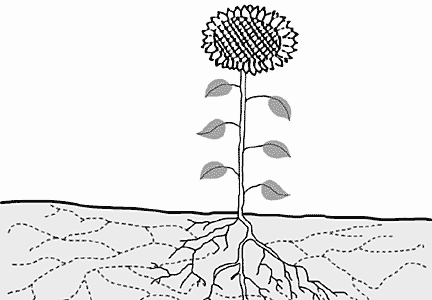
Схема, показывающая, как увеличивается площадь питания микоризованного растения.
Цветом изображены гифы гриба.Грибы могут быть причиной эволюционной сегрегации видов растений. Ржавчинные грибы из рода Uromyces в уредиальной (дикариотической) стадии поражают дикие виды ячменя Hordeum, а в гаплоидной — виды семейства лилейных Liliaceae. При экспериментальном заражении наиболее восприимчиво к ржавчине луковичное — Leopoldia eburnea. Она растет в пустыне Негев на Ближнем Востоке и цветет только в короткий период дождей. В остальное время даже не выходит на поверхность и никогда в природе не поражается ржавчиной. Полагают, что болезнь оттеснила Leopoldia как очень восприимчивый вид из генетического центра ячменя на периферию, где экологические условия неблагоприятны для ячменя и нет ржавчины.
Грибы защищают сложившийся фитоценоз от внедрения новых членов. Механизмы устойчивости к эндемичным болезням выработались как результат сопряженной эволюции. Виды растений, не эволюционирующие вместе с паразитами, попадая в новые районы, жестоко страдают от эндемичных болезней. Следовательно, эндемичные грибы ведут себя как рэкетиры, которые, снимая с растений некоторую долю фотосинтетической продукции, обеспечивают им «крышу».

Раковое поражение ствола каштана
(возбудитель болезни - гриб Criphonectria parasitica).
Веймутовая сосна (Pinus strobus),пораженная ржавчиной (возбудитель Cronartium ribicola).
Агроценозы
Итак, высший и низший организмы (растение и патогенный гриб) составляют целостную систему, в которой осуществляется взаимный отбор. Правила игры в ней таковы: оба вида находятся в равновесии лишь при определенных условиях. Их изменение может сдвинуть равновесие, чаще всего в пользу паразита. Колебания природных условий вызывают временные флуктуации, а антропогенные воздействия — приводят к необратимым последствиям. Такие изменения можно свести к четырем главным типам:
— пахотное земледелие с внесением удобрений резко меняет соотношение разных групп микроорганизмов в почве, давая преимущества в основном фитопатогенным видам. Снижается число микоризообразующих и сапротрофных грибов — антагонистов фитопаразитов;Как писал известный зоолог В.Л.Контримавичус, «конфликт между стабилизирующим экосистемы механизмом, каковым являются паразиты, и человеческой деятельностью — вероятно, наиболее драматический конфликт человечества с законами существования биосферы» [5]. В перспективе ожидается усиление этого конфликта, связанное с глобальным потеплением.— снижение фитоценологического и популяционного разнообразия растений в агроценозах по сравнению с природными ценозами обусловливает массовое накопление вирулентных штаммов фитопатогенов;
— завоз с растительным материалом новых патогенных грибов вызывает эпифитотии местных видов растений, не обладающих устойчивостью к ним. Так, гибель веймутовой сосны от ржавчины и каштанов от рака коры произошли вследствие вселения паразитов в новые районы и нападения на эндемичные растения;
— ухудшение условий жизни дикорастущих растений из-за накопления ксенобиотиков в воде, почве и воздухе, высокой рекреационной нагрузки и других факторов, связанных с деятельностью людей, делает их высоко чувствительными к патогенным грибам. Так, эвкалиптовые леса в Восточной Австралии погибли от корневой гнили, вызванной Phytophthora cinnamomi. От болезни страдали леса в местах с высокой рекреационной нагрузкой, где из-за уплотнения почвы и вытеснения воздуха корешки отмирали, открывая ворота инфекции.
Деструкция мертвых растений
Грибы, особенно гименомицеты, — единственная группа организмов, ферментативно разлагающая лигноцеллюлозный комплекс, из которого построены растения. Поэтому глобальная экологическая роль грибов лежит в сфере кругооборота углерода [6]. В России его годичное депонирование в древесные растения составляет 240—245 Мт C/год, а в древесину (стволы, ветки, корни) примерно 45 Мт С/год, т.е. 20—30%. Активное участие в балансе экосистемной и биомной продукции принимают грибы.
Нетто-экосистемная продукция — один из наименее изученных компонентов углеродного баланса (табл.1). Уравновешиваются ли потери углерода в ходе деструкции его накоплением в результате фотосинтеза и каковы временные соотношения между этими процессами? Окончательного ответа пока нет.
Углеродная емкость лесного детрита зависит от двух показателей: годичного поступления опада (листьев) и отпада (веток) — DD и скорости деструкции (табл.2). Листья разлагаются в полтора-два раза быстрее, чем древесина, что не удивительно, и кроме того, в холодном климате разрушение опада в четыре раза продолжительнее, чем в теплом [7]. В нашей стране, где наибольшие площади под лесами приходятся на холодную зону, ежегодно разлагается примерно 10% опада.
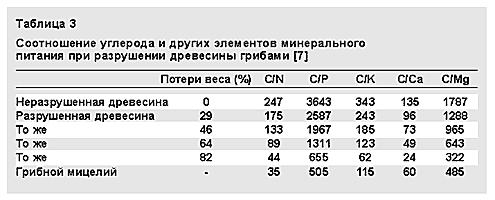
Как показали В.А.Мухин и Н.Т.Степанова, большая часть (до 80%) продуктов распада древесины превращается под действием грибов в углекислый газ и лишь 15—35% идет на построение мицелия [8]. Таким образом, грибы завершают углеродный цикл, начатый фотосинтезирующими растениями. Переход углерода в СО2 сопровождается освобождением других элементов, связанных в растительных клетках (табл.3).
За дальнейшие пути реализации углерода и других элементов питания ответственны организмы, которые питаются мицелием и плодовыми телами древоразрушающих сапротрофов (главным образом беспозвоночными).
Рассмотрим, наконец, нетто-биомную продукцию углерода, которая длительные сроки сохраняется в резервуаре (болотах — 113.5 Гт, почве — 235—366 Гт, древесине — 34.4 Гт). Наиболее подвижный углерод, хранящийся в почве (углерод гумуса), разлагается в результате микробиологических процессов, в которых активно участвуют и грибы. Поскольку их масса составляет примерно 90% от биомассы остальной почвенной микробиоты, соответствен и их вклад в разложение почвенного гумуса. Наименее подвижен углерод болот: его ежегодный импорт болотной растительностью составляет 20 г/м2, а эмиссия в виде углекислого газа и метана — 0.1—2.5 г. Поскольку в болотах преобладают анаэробные условия, основные деструкторы органического углерода не грибы, а бактерии.
Наконец, грибы, главным образом базидиальные, — единственные природные первичные деструкторы древесины. Процесс этот — очень медленный, зависит от толщины стволов и ветвей, видов грибов, климатических условий. Например, до полного разложения крупных стволов тсуги из семейства сосновых проходит около 3 тыс. лет, а мелких веток — от двух до 20 лет. В среднем релаксация углерода в древесине составляет 500—600 лет [9]. Быстрее всего связанный углерод освобождается при хозяйственном или случайном (в результате лесных пожаров) сжигании древесины.
Таким образом, люди в результате своей деятельности выводят из медленного оборота огромные массивы древесины, приводят их к быстрой минерализации, несравнимой с естественной скоростью связывания, создавая тем самым глобальные экологические проблемы.
Многообразные формы взаимоотношения грибов с растениями обусловили и их использование в биотехнологиях. Сегодня микоризация растений, способствующая повышению продуктивности и устойчивости к патогенным организмам — широко распространенный стандартный прием в сельском и лесном хозяйствах. Ферменты древоразрушающих грибов, утилизирующие лигнин, стали основой технологических процессов в древообрабатывающей промышленности многих стран. Контролирующие эти ферменты гены клонированы и перенесены в технологически удобные микроорганизмы (например, дрожжи). Нашли практическое применение даже фитопатогенные грибы. Споры некоторых видов накапливают в ферментерах и используют в качестве микогербицидов для опрыскивания засоренных посевов сельскохозяйственных культур. Несомненно, по мере изучения молекулярных механизмов фитопаразитизма горизонты практического применения грибов будут расширяться. * * *
Литература
1. Strackle S., Kisner K. et al. // Nature. 2002. V.417. P.959—962.
2. Simard S.W., Perry D.A. et al. // Nature. 1997. V.388. P.579—582.
3. Беклемишев И.Н. Возбудители болезней как члены биоценозов // Биоценолог. основы сравн.
паразитологии. М., 1970. С.334—352.4. Packer A., Clay K. et al. // Nature. 2000. V.404. P.278—281.
5. Контримавичус В.Л. // Журн. общ. биологии. 1982. Т.423. Вып.3. С.292—302.
6. Кругооборот углерода на территории России / Под ред. Г.А.Заварзина. М., 1999.
7. Boddy L., Watkinson S.C. // Can. J. Bot. 1995. V.73. P.1377—1383.
8. Мухин В.А., Степанова Н.Т. // Экология. 1976. №6. С.42—45.
9. Исаев А.С., Коровин Г.Н. Углерод в лесах Северной Евразии // Кругооборот углерода на территории России. М., 1999. С.63—95.
Апрель 2003