
№ 4, 2007 г.
© Малярчук Б.А., Деренко М.В.
Структура русского генофонда
Б.А.Малярчук, М.В.Деренко
Борис Аркадьевич Малярчук, д. б. н., зав. лаб. генетики Ин-та биологических проблем Севера Дальневосточного отделения РАН (Магадан).
Мирослава Васильевна Деренко, к. б. н., вед. научн. сотр. того же института.Немного славянской истории
Прежде чем рассказать о результатах исследования структуры русского генофонда, напомним - буквально в нескольких словах - историю славян. Эту самую многочисленную группу народов в Европе принято делить на западных славян, южных и восточных. К западным относят поляков, чехов, словаков и лужичан; к южным - болгар, сербов, хорватов, словенцев, македонцев, боснийцев, черногорцев, а к восточной - русских, украинцев и белорусов. Вполне естественно, что вопросами происхождения славян занимаются многие ученые разных специальностей: историки, археологи, языковеды, этнографы, антропологи и др. Делаются попытки восстановить этническую историю народов по предметам материальной культуры, на основании языка и письменности, по физическим типам людей.
Совокупность полученных данных приводит исследователей к разным гипотезам, которые можно разделить на две диаметрально противоположные по смыслу группы. По гипотезам одной группы, славянские народы произошли от общих предков; в соответствии с гипотезами другой группы, предки у нынешних славянских народов были разные, т.е. они не связаны общностью происхождения.
Сейчас в славянской группе народов, как, впрочем, и любых других, виден результат их длительной истории. Поэтому и не удивительно, что современные славяне антропологически неоднородны и даже принадлежат к разным ветвям европеоидной расы. Признаки, характеризующие отдельные славянские группы, проявляются и у соседних народов. Причину тому исследователи видят в расселении древних славян на обширных территориях Европы, интенсивнее всего проходившем в VI-VII вв. [1]. В новых местах они контактировали с разными народами: с германцами на западе, с фракийцами, кельтами, иллирийцами на юге, с иранцами и финно-уграми на востоке, с балтами на севере. Естественно, что инородцы воздействовали на славян и со временем внесли собственные черты в их антропологический облик.
По археологическим и лингвистическим данным, прародиной славянских народов был Северокарпатский регион, включавший Галицию, Волынь и Подолье, откуда и началось продвижение по Европе. В VII-VIII вв., по мнению В.О.Ключевского, славяне восточной ветви, занимавшие северо-восточные склоны Карпат, стали продвигаться на восток и северо-восток Европы под натиском аваров [2].
Уже ко времени переселения славян в Восточную Европу соотношение исходных антропологических компонентов в их группах могло отличаться. По мере же освоения восточноевропейских территорий в разных направлениях все более проявлялся полиморфизм (разнообразие сочетаний антропологических признаков), обусловленный смешением с местным населением. Например, по комплексу расоводиагностических признаков белорусы и русские сходны с северо-западными народами (т.е. сказывается финно-угорское и летто-литовское влияние), а украинцы - с южными (из-за иранского и романского влияния) [3].
Становление государственности на Руси, очевидно, в немалой степени способствовало межэтническим взаимодействиям в пределах европеоидной расы. Свой вклад в метисацию внесло и освоение новых земель на востоке Евразии, где славянское население соприкасалось с монголоидным. В Восточной Европе Русь расширялась за счет колонизации северо-восточных территорий; там строились русские города и заставы, заселяемые восточными славянами "из разных мест, из разных племен". Поэтому славянское население Руси издревле различалось антропологически. Продвигаясь из Руси древней, юго-западной, в Русь новую, северо-восточную, оно смешивалось с автохтонным балтским и финно-угорским населением.
Историки полагают, что в Восточной Европе до IX в. существовали два территориальных массива славян - юго-западный и псковско-новгородский, объединение которых привело к созданию Древнерусского государства [4]. Таким образом, исторические сведения, археологические и лингвистические данные свидетельствуют об антропологической гетерогенности исходного населения, ставшего основой русского народа. Действительно, обобщенный картографический анализ антропологических данных выявляет в составе русского населения три "ядерные структуры" - восточную, центральную и западную. Первая сформировалась на основе дославянского субстрата, вторая - за счет длительной метисации, третья - за счет исходного заселения славянским народом [5].
Такой, в весьма кратком изложении, вырисовывается история славянских народов, и русского в том числе, на основе археологических, лингвистических и антропологических сведений. Подтверждается она, подвергается исправлениям или вовсе отвергается результатами генетических исследований, активно проводимых в последние десятилетия? Чтобы ответить на этот вопрос, приведем здесь собственные сведения по молекулярной антропологии русского народа.
Родительские линии ДНК в генофонде русских
Этническое своеобразие любого народа обусловлено генетическими процессами, которые сиюминутно внешне себя ничем не проявляют. Но о них можно узнать, оценив сходства и отличия в детальном строении ДНК - митохондриальной (мтДНК), которая, напомним, наследуется по материнской линии, и Y-хромосомы, наследуемой по отцовской линии. Сейчас в мире уже существуют богатые базы данных, содержащие сведения о последовательности нуклеотидов в этих молекулах, на их основе строятся филогенетические древа. Относительно недавно, в последние 10-20 лет, возникла даже самостоятельная научная дисциплина - молекулярная антропология. Ее объектами и служат мтДНК и Y-хромосома, а конечной целью - генетическая история того или иного народа. Число исследований такого рода растет с каждым годом, мы тоже применяем молекулярно-генетический подход для изучения структуры геномов разных этносов, чтобы узнать их истоки [6].
Реконструировать процессы, которые приводят к формированию генофондов популяций, этнических групп и этнорасовых общностей, выяснить, как они распределялись пространственно за время их истории, можно с использованием больших массивов данных об изменчивости ДНК. Что касается евразийских народов, то в современный анализ филогеографической изменчивости * ДНК вовлечены много тысяч человек из различных стран и регионов. По этим вполне представительным статистическим выборкам в линии мтДНК удалось выделить монофилетические группы и подгруппы, а затем реконструировать последовательность ее эволюционных изменений.
* Филогеографическая изменчивость - это распределение вариантов ДНК этноса (популяции, группы) за время существования в определенном географическом пространстве.На сегодня уже установлено, какие митохондриальные линии присущи населению Западной и Восточной Евразии, Африки, Австралии и Америки. По современным представлениям, все евразийские группы мтДНК входят в состав трех макрогрупп - M, N и R, которые произошли примерно 65 тыс. лет назад из африканской митохондриальной группы L3. Важно, что распределение линий мтДНК в генофонде населения Евразии характеризуется выраженной этнорасовой специфичностью. Например, генофонды народов западной и восточной частей Евразии различаются кардинально. Почти все линии мтДНК корейцев и бурят относятся к набору восточноевразийских групп - A, B, C, D, F, G, M7, M8a, M9, M10, M11, Y, Z, N9a, R9. Европейцам же свойственны группы H, HV*, pre-V, T, J, K, U2, U3, U4, U5, U8, N1a, I, W и X. В другой части Евразии, в северной, судя по полученным к настоящему времени данным, лишь в краевых популяциях почти полностью преобладают митохондриальные линии одного этнорасового происхождения - монголоидного или европеоидного. Огромная же по протяженности территория (от Алтае-Саянского нагорья до Восточно-Европейской равнины) представляет собой древнюю контактную зону. Именно в ней проходил расогенез за счет межрасового смешения.

Филогенетическое дерево митохондриальной ДНК человека.
В начале 90-х годов мы занялись исследованием этнической истории русского народа, для чего изучали вариабельность материнских и отцовских линий ДНК, т.е. мтДНК и Y-хромосомы. Чтобы выявить изменчивость мтДНК в популяциях, использовали комбинированный прием: анализировали рестрикционный полиморфизм * фрагментов кодирующих районов и изменчивость нуклеотидных последовательностей некодирующих гипервариабельных участков. В результате выяснилось, что митохондриальный генофонд русского населения характеризуется высоким разнообразием, но встречались и общие группы. Наиболее частыми из них были H, U, T и J - те же группы, которые широко распространены в генофондах других европейских народов. Монголоидная примесь у русских оказалась незначительной - около 1.5%, причем ее составляли группы мтДНК (C, D, M*, G2a, N9a) восточноевразийского происхождения.
* Имеются в виду различия в наборах участков мтДНК, возникающих при ее расщеплении (рестрикции) специальными ферментами. В данном случае расщеплению подвергали не всю мтДНК, а только кодирующие участки.Необходимо было попытаться выяснить истоки разнообразия мтДНК у русских, понять, не следствие ли это особенностей формирования славянского этноса. Поэтому мы посмотрели, как распределяются гаплотипы мтДНК в европейских популяциях: русских, поляков, боснийцев, словенцев, финнов, французов, немцев. Судя по результатам этого филогеографического анализа, митохондриальные генофонды европейцев имеют общие черты. Маркеры же, которыми одни этнические группы и общности отличаются от других, представлены, как правило, комбинациями редких подгрупп и отдельных гаплотипов мтДНК. Следовательно, существует единый генетический субстрат, на основе которого развивались генофонды западных и восточных славян, а также их соседей - германцев и западнофинских народов (эстонцев и финнов) [7]. Отдельные митохондриальные линии этого субстрата попали и в генофонд народов, населявших юг Европы и Балканы (по-видимому, в основном в процессе славянских миграций). Об этом можно судить, приняв во внимание тот факт, что южнославянские популяции отличаются генетически от их соседей - греков, албанцев и итальянцев [8].Основываясь на данных об изменчивости мтДНК, мы попытались выяснить, что стало с этой молекулой при дифференциации славян на восточных, западных и южных [9]. Кроме того, посмотрели, как сказалось такое разделение на генофондах соседних популяций Европы и Западной Азии. Оказалось, что между разными группами славян существуют не только антропологические, но и генетические различия. Их сила определяется в значительной мере степенью метисации с тем дославянским населением, которое существовало в пределах современного этнического ареала славян. Кроме того, сказывается и интенсивность, с которой они взаимодействовали с соседними народами. Последний вывод следует из того, что соседи русских, западнофинские популяции, характеризуются относительно высоким генетическим сходством с русскими, германские популяции - с западными славянами, а балканские - с южными. Проще говоря, в молекулах мтДНК соседей запечатлены характерные славянские черты - признак тесного взаимодействия пришлых и местных народов. Из всех этих результатов напрашивается вывод о единстве происхождения славян, центральное положение среди которых принадлежит западным группам. Иначе говоря, генофонд западных славян включает максимальное число типов мтДНК, встречающихся как у восточных, так и у южных славян, хотя между этими двумя группами нет высокого генетического сходства. По всей видимости, можно предположить, что генофонд западных славян ближе к исконно славянскому, но это пока всего лишь предположение.
Другая генетическая система, изменчивость которой теперь активно изучается в филогеографических исследованиях, - это нерекомбинирующая часть Y-хромосомы. С ее помощью можно составить представления о распространенности в популяциях тех или иных маркеров, которые, как и в мтДНК, несут информацию о генетической истории этнорасовых групп человека.
В своих исследованиях мы тоже анализировали изменчивость Y-хромосомы, изучая разнообразие ее линий в русских популяциях. Для этого использовали биаллельные SNP-локусы * ее ДНК, так как вариантами их полиморфизма определяется топология филогенетического дерева Y-хромосомы. Было проведено исследование около 400 индивидуальных препаратов ДНК русского населения из восьми областей - Белгородской, Орловской, Тульской, Калужской, Владимирской, Нижегородской, Ярославской и Псковской.
* SNP (Single Nucleotide Polymorphism - однонуклеотидный полиморфизм)-локусы - это участки ДНК, которые отличаются один от другого единичной нуклеотидной заменой, например: GTAC и GCAC.В результате выяснилось, что мужской генофонд русских представлен 14 группами Y-хромосомы (P*, R1*, R1a, N*, N3, BR*, C, F*, G, I, J, H, E, K*). Однако большинство линий (80%) относятся лишь к трем из них - R1a, I и N3. Наиболее интересными представляются группы R1a и N3, поскольку в Европе первая маркирует славянские народы, а вторая распространена с наибольшей частотой (более 50%) у финно-угорского и балтского населения Северной и Восточной Европы и практически отсутствует в генофондах западных и южных европейцев, включая славян [10]. Там, где славяне соседствуют с другими этническими общностями, в ДНК Y-хромосомы последних резко снижается частота встречаемости R1a [10, 11]. Так, например, характеризуются немцы, живущие в приграничных с поляками или с сорбами (лужицкими сорбами, или лужичанами) районах.В генофонде русских частота встречаемости N3 варьирует в широком диапазоне: от 5% в Калужской обл. до 35% в Псковской. Это свидетельствует об отличающейся степени взаимодействия между славянским и финно-угорским/балтским населением в разных частях восточноевропейского ареала русских. Из проанализированных нами образцов Y-хромосомы лишь в псковской и поморской (этнографической группы русского населения Архангельской обл.) популяциях частота гаплогруппы N3 оказалась сопоставимой с той, что характеризует финно-угров и балтов [12].
Примечательно, что более тонкими исследованиями полиморфизма Y-хромосомы можно различить две группы в N3 - "балтскую" и "финно-угорскую" [13]. Нам удалось это сделать по результатам анализа 12 STR-локусов *, которые эволюционируют с более высокой скоростью, чем упомянутые биаллельные SNP-локусы. Оказалось, что высокая частота гаплогруппы N3 в псковской популяции обусловлена преимущественно "балтскими" вариантами Y-хромосомы (их доля составляет 64%). В генофондах русского населения центральных и южных областей европейской части России преобладают (в среднем до 80%) N3-варианты Y-хромосомы, характерные для финно-угорского населения Восточной Европы.
* STR-(Short Tandem Repeats - короткие тандемные повторы)-локусы - это относительно короткие (до 6 пар нуклеотидов) участки ДНК, расположенные в молекуле в виде блоков из повторяющихся последовательностей. Иначе STR-локусы называют микросателлитными последовательностями ДНК.Итак, у славянских народов набор маркеров в ДНК Y-хромосомы сходен. Но, что важно, существующие отличия славян от соседей (германцев на западе Европы; греков, албанцев и итальянцев на юге; финно-угорского населения на востоке Европы) по Y-хромосоме гораздо заметнее, чем по линиям мтДНК [10, 12].Дифференциация родительских линий ДНК
Наиболее информативен для исследований судьбы этносов анализ генетической дифференциации популяций по данным об изменчивости мтДНК и Y-хромосомы. Результаты такого рода анализа позволяют составить представление о том, как распространялись материнские и отцовские линии ДНК в процессе экспансии народов. Хороший пример в этом отношении - русские, освоившие за последние почти полторы тысячи лет обширные пространства Евразии.
Мы изучили мтДНК и Y-хромосомы в 10 популяциях русского населения Восточной Европы: от Ставропольского края на юге до Псковской обл. на севере и от Орловской обл. на западе до Нижегородской на востоке. Кроме собственных результатов, мы привлекли в анализ данные о поморской, костромской, ростовской и курской популяциям, полученные другими научными коллективами.
В итоге выяснилось, что по отцовским линиям русское население отличается в значительно большей степени, чем по материнским. Так, коэффициенты дифференциации FST * Y-хромосомы составляют 3.1%, в то время как мтДНК - лишь 0.42%, т.е. различаются почти на порядок. Примечательно в этих коэффициентах еще одно: по Y-хромосоме у русских они существенно ниже, чем в Европе в целом (там их величина достигает 7.04%), а по митохондриальным линиям не отличаются (0.42% и 0.41%).
* FST - это коэффициент локальной генетической дифференциации, мера генетической подразделенности популяции и одновременно эквивалент инбридинга субпопуляции (S) относительно всей популяции (T).Хотя в популяциях русского населения различия в распределении материнских линий ДНК не очень высоки, статистическим анализом нам удалось выявить две ветви - юго-западную и северо-восточную. Каждая из них образует кластер с другими европейскими популяциями. Так, юго-западная ветвь мтДНК входит в кластер материнских линий, свойственных народам Центральной Европы, - как западнославянским (например, полякам), так и балтским (литовцам), а также некоторым финно-угорским (эстонцам). Северо-восточная ветвь мтДНК русских проявляет сходство с мтДНК финно-угорских и других народов Восточной Европы (финнов, карелов, марийцев, татар, адыгейцев).Результаты анализа FST-дистаций, полученных на основании полиморфизма биаллельных локусов Y-хромосомы, также свидетельствуют о гетерогенности русских популяций. Однако картины дифференциации по мтДНК и Y-хромосоме не совпадают. В сравнительном анализе использовались литературные данные [10] об изменчивости Y-хромосомы украинцев, белорусов, литовцев, латышей, эстонцев, финнов, марийцев и чувашей, т.е. соседних с русскими этнических групп. Собственные сведения о полиморфизме Y-хромосомы русского населения в Восточной Европе мы дополнили также данными об изменчивости этой хромосомы у поморов [14]. Из анализа всей совокупности ее характеристик следует, что лишь псковская и поморская популяции проявляют выраженное сходство с финно-угорскими и балтскими народами Северной и Восточной Европы. Подавляющее же большинство русских популяций по особенностям генетических структур образуют единый кластер вместе с такими славянскими народами, как поляки, украинцы и белорусы. У всех у них высока частота группы R1a, а группа N3 встречается с умеренной или даже низкой частотой. Эти же группы Y-хромосомы в псковской и поморской популяциях встречаются в обратной пропорции (как у финно-угорских и балтских народов): частота N3 повышена, а R1a - понижена.
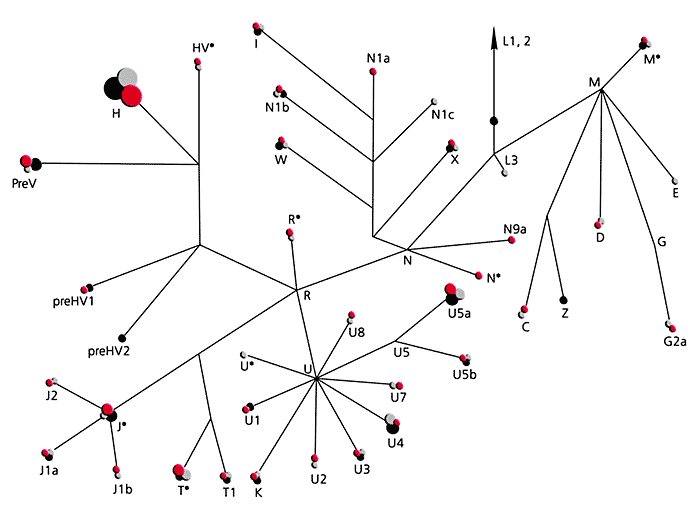
Структура митохондриального генофонда русских и еще двух славянских народов - поляков и боснийцев.
Кластеры (группы и подгруппы) мтДНК обозначены буквами;
розовыми кружками (их размер пропорционален частоте гаплотипа мтДНК) даны кластеры
русских, серыми - поляков, черными - боснийцев.Таким образом, результаты комбинированного анализа позволяют заключить, что в русских популяциях центральноевропейский генетический компонент как материнского, так и отцовского происхождения выражен по-разному. Этот компонент преобладает лишь в южной и западной частях восточноевропейского ареала русских. В центре и на востоке ареала начинают доминировать восточноевропейские линии мтДНК. И только в некоторых популяциях северной части ареала русских комбинация линий мтДНК и Y-хромосомы та же, что у финно-угорского населения. Выявленная картина генетической дифференциации современных русских, по-видимому, отражает процессы, обусловленные межэтническим взаимодействием в ходе формирования русского народа начиная с эпохи средневековья.
Итак, мы выяснили молекулярно-генетическую структуру русского генофонда, правда, пока лишь в считанном количестве популяций. В целом она согласуется с результатами картографического анализа антропологических данных, ставшими основой гипотезы об истории формирования русского народа. Судя по всему, лишь на западе этнической территории русские представляют собой генетических потомков славян, в центральной части русский этнос сформировался за счет смешения славян и дославянского финно-угорского населения, а на востоке и в северной части ареала произошло только замещение дославянских языков и культуры славянскими [5]. Между тем молекулярно-генетические данные позволили глубже понять причины наблюдаемых генетических и антропологических различий. Исходя из распределения линий мтДНК и Y-хромосомы в русских популяциях, становится очевидным, что наблюдаемая "трехзональная" картина их дифференциации обусловлена различным участием мужчин и женщин в освоении восточноевропейских территорий. По всей видимости, лишь начальный этап колонизации осуществлялся как мужчинами, так и женщинами (видимо, племенными группами, родами, семьями), а более поздние - преимущественно мужчинами, которые женились на аборигенках.
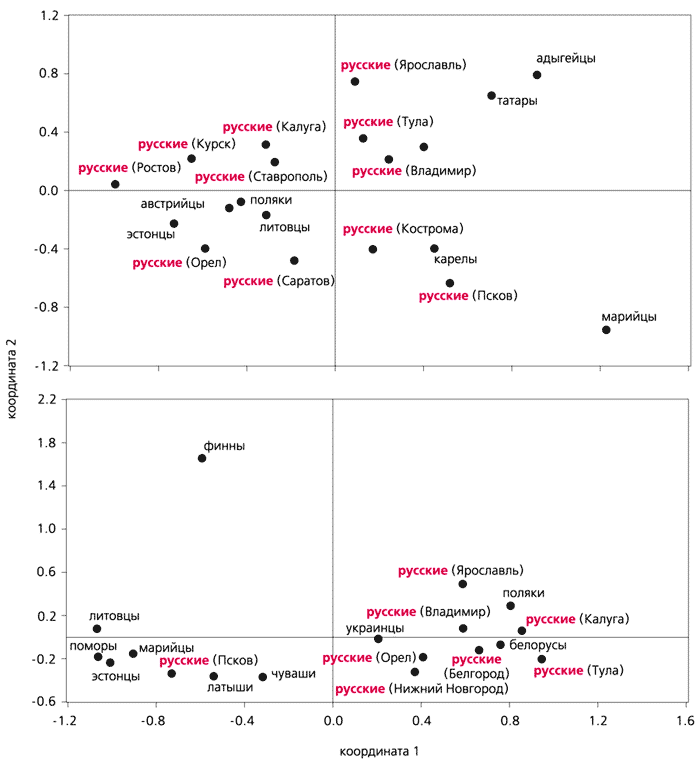
Расположение европейских популяций в пространстве двух координат.
"Карты" построены на основании результатов многомерного шкалирования (одного из методов многомерной статистики) межпопуляционных FST-дистанций по данным об изменчивости мтДНК (вверху) и Y-хромосомы. Для анализа использованы также литературные данные [10, 19, 20]. Видно, что по мтДНК русские, живущие в разных областях, не очень значительно отличаются друг от друга, но все же разделяются на юго-западную и северо-восточную зоны. В первой, кроме русских, оказываются некоторые народы из западных славян (поляки), балтов (литовцы) и финно-угров (эстонцы). Во вторую зону вместе с русскими популяциями попадают несколько финно-угорских народов, а также татары и адыгейцы. Расположение популяций по Y-хромосоме отражает своеобразие их мужского генофонда, определяемого, главным образом, высокой частотой группы N3 (61%) и очень низкой (10%) - R1a. Если сравнить обе "карты", становятся очевидными генетические различия между генофондами популяций по мтДНК и Y-хромосоме. Особенно это касается финнов: в первом случае они располагаются среди русских Ярославской, Владимирской и Тульской областей, а по Y-хромосоме занимают обособленное положение [10].* * * Полученная в нашей работе картина генетической дифференциации русского населения Восточной Европы нуждается, конечно, в уточнениях. В связи с этим нужно изучить в дальнейшем изменчивость мтДНК и Y-хромосомы в тех русских популяциях, которые еще не охвачены такими исследованиями. Кроме того, необходимо узнать, каким было генетическое разнообразие древнего восточноевропейского населения в разные периоды его истории, начиная от дославянского.Как показывают результаты анализа ДНК, генетическая преемственность между древними и современными народами соблюдается отнюдь не всегда, ведь одни народы могли сменяться другими. Например, генофонды неолитического и нынешнего населения Прибайкалья имеют мало общего [15]. На возможную смену этносов в некоторых европейских регионах - Тоскании (Италия) и Стране басков (Испания) - указывают результаты генетических исследований древнего и современного населения [16]. Более того, уже имеются примеры существенных противоречий между наблюдаемыми и ожидаемыми распределениями маркеров мтДНК в палеопопуляциях. Так, у неолитического населения Страны басков не обнаружена митохондриальная группа V. А ведь именно ей придается большое значение, когда, основываясь на распределении групп мтДНК у современных европейцев, моделируют процесс послеледникового заселения Европы выходцами из пиренейского "убежища" [17].
Выявлено и еще одно серьезное несоответствие. У неолитического населения Центральной Европы, ведущего происхождение от "первых европейских фермеров" (носителей переднеазиатской технологии агрокультуры), митохондриальная группа N1a встречается с неожиданно высокой частотой. Современным же европейцам эта группа практически не свойственна и потому никак нельзя было предсказать ее высокую распространенность среди неолитического населения [18].
Приведенные примеры показывают, насколько важна проверка этноисторических моделей, если выявленное генетическое разнообразие современных народов переносится в их прошлое. Выходит, экстраполяции не всегда надежны. Каким был генофонд далеких предков, безошибочно можно судить по результатам исследований ДНК непосредственно древнего населения. И такой анализ вполне возможен, ведь археологами найдено множество останков неолитического времени, пригодных для выделения ДНК.
Работа выполнена при финансовой поддержке Российского фонда фундаментальных исследований (проекты 00-06-80448, 03-04-48162).
Литература
1. Алексеева Т.И., Алексеев В.П. Антропология о происхождении славян // Природа. 1989. №1. С.60-69.
2. Ключевский В.О. Сочинения: В 9 т. Т. I. Курс русской истории. Ч.I. М., 1987.
3. Алексеева Т.И. Этногенез восточных славян (по данным антропологии). М., 1973.
4. Янин В.Л., Алешковский М.Х. // История СССР. 1971. №2. С.32-61.
5. Рычков Ю.Г., Балановская Е.В. // Вопр. антропол. 1988. Вып.80. С.3-37.
6. Деренко М.В., Малярчук Б.А. Генетическая история коренного населения Северной Азии // Природа. 2002. №10. С.69-76.
7. Malyarchuk B.A., Grzybowski T., Derenko M.V. et al. // Ann. Hum. Genet. 2002. V.66. P.261-283.
8. Malyarchuk B.A., Grzybowski T., Derenko M.V. et al. // Ann. Hum. Genet. 2003. V.67. P.412-425.
9. Малярчук Б.А. // Генетика. 2001. Т.37. №12. С.1705-1712.
10. Rosser Z.H., Zerjal T., Hurles M. et al. // Am. J. Hum. Genet. 2000. V.67. P.1526-1543.
11. Behar D.M., Thomas M.G., Skorecki R. et al. // Am. J. Hum. Genet. 2003. V.73. P.768-779.
12. Malyarchuk B., Derenko M., Grzybowski T. et al. // Hum. Biol. 2004. V.76. P.877-900.
13. Zerjal T., Beckman L., Beckman G. et al. // Mol. Biol. Evol. 2001. V.18. P.1077-1087.
14. Wells R.S., Yuldasheva N., Ruzibakiev R. et al. // Proc. Natl. Acad. Sci. USA. 2001. V.98. P.10244-10249.
15. Mooder K.P., Schurr T.G., Bamforth F.J. et al. // Am. J. Phys. Anthropol. 2006. V.129. P.349-361.
16. Alzualde A., Izaguirre N., Alonso S. et al. // Ann. Hum. Genet. 2005. V.69. P.665-679.
17. Torroni A., Bandelt H. -J., D’Urbano L. et al. // Am. J. Hum. Genet. 1998. V.62. P.1137-1152.
18. Haak W., Forster P., Bramanti B. et al. // Science. 2005. V.310. P.1016-1018.
19. Orekhov V., Poltoraus A., Zhivotovsky L.A. et al. // FEBS Letters. 1999. V.445. P.197-201.
20. Richards M., Macaulay V., Hickey E. et al. // Am. J. Hum. Genet. 2000. V.67. P.1251-1276.
