№4, 2006 г.
© Павлинов И.Я.
На пути к филогенезу
И.Я. Павлинов
Игорь Яковлевич Павлинов, д.б.н., вед.н.с. Зоологического музея МГУ.
Когда смотришь сзади на уходящий поезд, виден только последний вагон - и не видно локомотива, тянущего за собой весь состав. В науке этот “эффект последнего вагона” означает, что каждый новый подход стремится заполнить собой всю познавательную ситуацию, претендуя на окончательное решение извечных проблем.
В современной филогенетике таким “последним вагоном” служит молекулярный подход, стремящийся приписать себе доминирующую роль. Однако при этом упускается из виду, что работать приходится с теми же задачами и концепциями, которые сложились в пору формирования классической биологии. Переоценка новизны и недооценка преемственности в развитии современной филогенетики порождает избыточный оптимизм в отношении новых способов решения старых задач.
* * *
В современной биологии значение эволюционной идеи велико, как ни в каком другом разделе естествознания. Причина в том, что материал по разнообразию живых организмов дает больше всего пищи для размышлений о превращениях одних вещей в другие. И вряд ли случайно, что формирование современного глобального эволюционизма началось именно с дарвиновской теории эволюции, объясняющей происхождение биологических видов.
Поскольку биологическое разнообразие есть результат длительного исторического развития, то понять строение и функционирование живых существ невозможно без знания их истории. Это обстоятельство делает исторические реконструкции одной из приоритетных задач в современной биологии.
Познанием путей и закономерности исторического развития организмов занимается особая дисциплина - филогенетика. На протяжении своей истории она знала времена и взлета, и падения, связанные как с успехами самой биологии, так и с развитием представлений о том, что такое научное знание. В последней трети XX в. интерес к историческим реконструкциям заметно возрос: биология столкнулась с тем же весьма специфическим феноменом, что и в конце XIX в., название которому - “филогенетический бум”.
Филогенез и филогенетика
Филогенетика зародилась в 60-е годы XIX в. вскоре после выхода в 1859 г. книги Ч.Дарвина “Происхождение видов путем естественного отбора”. Термин “филогенез” впервые появился в 1866 г. в фундаментальной работе немецкого биолога-эволюциониста Э.Геккеля “Общая морфология животных”. Этим понятием автор обозначил и процесс исторического развития организмов, и структуру родственных (филогенетических) отношений между ними. Введенный приблизительно в эти же годы английским философом Г.Спенсером в научный оборот термин “эволюция” в его современном историческом понимании (ранее им обозначали индивидуальное развитие) также быстро завоевал популярность.
В результате понятия филогенеза и эволюции стали восприниматься как очень близкие по смыслу. Такая классическая трактовка бытует и по сей день: филогенез определяют как пути, закономерности и причины исторического развития организмов [1]. Выдающийся отечественный эволюционист И.И.Шмальгаузен рассматривал филогенез как цепочку сменяющих друг друга онтогенезов.
В настоящее время формируется биотоцентрическая концепция филогенеза, согласно которой биологическая эволюция - это саморазвитие биоты как целостной системы, а филогенез - один из аспектов этого развития [2]. Такое понимание биологической эволюции вообще и филогенеза в частности более всего соответствует современным представлениям об общих законах развития, которые разрабатывает синергетика [3].
Один из важнейших результатов эволюции - глобальная структура биоты Земли, которая проявляется в многоуровневой иерархии групп, по-разному интегрированных и организованных. В некотором приближении можно считать, что эта структура состоит из двух иерархий. Одна из них связана с разнообразием биоценозов (природных экосистем), объединенных экологическими отношениями. Историческое становление этой иерархии обозначают как филоценогенез [4]. Вторая связана с разнообразием таксонов - филогенетических групп, члены которых находятся в родственных отношениях. Становление именно этой иерархии и есть филогенез [2, 5].
В свою очередь, в нем выделяется несколько компонент, позволяющих исследовать историческое развитие с разных сторон. В начале XX в. австрийский палеонтолог О.Абель разграничил их следующим образом: ряды предков - “истинные филогенезы”; ряды приспособлений, касающихся одного органа; ряды ступеней совершенствования организации. В настоящее время “истинный филогенез” - развитие (появление и/или изменение состава) филогенетических групп организмов как таковых, безотносительно их свойств, - обозначают как кладогенез, или кладистическая история. Этот термин предложил английский биолог Дж.Гексли в 40-х годах, противопоставив его анагенезу - изменению уровня организации живых существ в процессе эволюции. Исторические изменения отдельных органов и свойств организмов немецкий систематик и эволюционист В.Циммерман в 60-х годах предложил называть семофилогенезом. Более удачен термин семогенез, обозначающий историческое развитие (появление, изменение, исчезновение) отдельных морфологических и иных структур, рассматриваемые вне связи с конкретными группами организмов, которым они присущи.
Группы, порождаемые кладогенезом, называют кладами: например, хордовые, а в их пределах - позвоночные. Группы, порождаемые анагенезом, - грады, ступени эволюционного развития: таковы многоклеточные животные по отношению к одноклеточным, а среди позвоночных - гомойотермные животные (птицы и млекопитающие) по отношению к пойкилотермным (низшие позвоночные). Принципиальное отличие двух этих категорий состоит в том, каким путем приобретаются общие свойства. Клады наследуют их от общего предка, а у град они появляются а результате параллельной или конвергентной эволюции. Современная филогенетика по преимуществу изучает становление иерархии филогенетических групп и специфичных для них свойств, ее основная задача - реконструкция кладогенеза. Из этого видно, что на нынешнем этапе своего развития филогенетика является по преимуществу кладогенетикой.
От естественной системы к эволюционной идее
В классической биологии в XVII-XIX вв. одним из ключевых было представление о естественной системе как всеобщем законе, которому подчинено все сущее. Эта система объединяет естественные группы организмов, существующие в самой природе. Ее воссозданием занималась и занимается систематика, задача которой состоит в том, чтобы распознать эти группы по их “естеству”.
Коль скоро конечная цель научного познания - выяснение причин того или иного явления, сама естественная система нуждалась в объяснении: в чем ее изначальная причина. В Средние века таковой считался божественный план творения. Со становлением науки Нового времени причину стали искать в самой Природе: так в естественнонаучное мировоззрение вошла идея исторического развития, давшая наиболее правдоподобное с материалистической точки зрения толкование естественной системы.
Важнейшую роль в формировании систематики и филогенетики сыграла немецкая натурфилософия. С ней связано, с одной стороны, восходящее к Аристотелю представление о “Великой лестнице Природы” (Г.Лейбниц), с другой - уподобление Природы сверхорганизму (Ф.Шеллинг, Л.Окен). Они дали два способа представления естественной системы - линейную и иерархическую. В первом случае имелся в виду непрерывный ряд организмов (“природа не делает скачков”), направленный от низших форм жизни к высшим. Во втором случае подразумевалось, что биота, подобно организму, поделена на части разного уровня общности - естественные группировки.
Уподобление биоты сверхорганизму имело особое значение для формирования эволюционного мировоззрения: ведь живое существо немыслимо без развития, направленного в сторону все большего совершенства и дифференциации. На этой основе, вкупе с идеей “лестницы совершенствования” (другое название “лестницы природы”), сформировалась ключевая идея классического эволюционизма, а с ним и классической филогенетики - уподобление исторического развития биоты индивидуальному развитию организма.
Эволюционная интерпретация непрерывного ряда форм породила учение Ж.-Б.Ламарка, согласно которому основная задача эволюционной теории сводилась к объяснению того, каким образом возникают новые свойства организмов, обеспечивающие повышение их уровня организации. Так в филогенетике возникла адаптационистская парадигма.
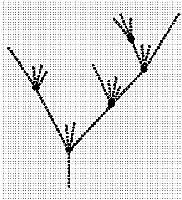
Иерархической системе более всего соответствует дивергентная концепция эволюции, связываемая с именем Ч.Дарвина. В данном случае история фактически сведена к многократно ветвящимся “цепочкам” превращения одних видов в другие (рис.1). Такое видение эволюционного развития сформировало в филогенетике генеалогическую парадигму.
Рис.1. Теоретическая схема генеалогических отношений между видами по Дарвину. Биологи с натурфилософским складом ума воспринимали историческое развитие как макромасштабный процесс. Для них сверхзадачей была (и по сей день остается) реконструкция становления основных групп живых организмов - царств, типов, ступеней развития. Для сторонников набиравшей силу дарвиновской трактовки эволюции ключевыми были события, происходящие на видовом уровне. Соответственно, в реконструкции истории основной акцент делается на “локальные” события - превращения одних видов в другие.
В начале XX в. вся натурфилософская картина мира была признана “ненаучной”, поскольку ее утверждения не проверялись опытным путем. В новое мировоззрение как нельзя лучше вписался дарвиновский селекционизм, дополненный мутационной теорией. Постепенно сложилась синтетическая теория эволюции, идеологическую основу которой составило популяционное мышление. В ней терминологически закрепилось разделение исторического развития на микроэволюцию и макроэволюцию (автор терминов - Ю.А.Филипченко); при этом была абсолютизирована первая и фактически отвергнута вторая.
Сведение механизмов эволюции к популяционным процессам позволило рассматривать филогенез как их “механическое” следствие, возникающее просто из-за того, что жизнь существует и развивается очень продолжительное время. Такое развитие эволюционной идеи в первой половине - середине XX в. сделало филогенетику в известной степени “вещью второго сорта”, лишило ее прежнего приоритетного статуса. Впрочем, во второй половине того же столетия популяционные идеи сыграли значительную роль в формировании современного этапа развития этой дисциплины.
Классическая филогенетика
Понимание филогенеза как закономерного исторического процесса не мешает по-разному трактовать смысл филогенетического единства, выделяющего естественные группы. Это породило несколько линий развития классической филогенетики, которые соответствуют трем уже упоминавшимся ключевым компонентам филогенеза - кладогенезу, семогенезу и отчасти анагенезу.
Признание филогенетического единства преимущественно как общности происхождения, т.е. монофилии, стимулировало развитие филогенетики в кладогенетическом направлении. Именно для кладогенетики своего рода символом стала метафора ветвящегося филогенетического дерева с единственным корнем.
В геккелевой трактовке эта концепция филогенеза соединяет натурфилософское и дарвиновское понимания эволюции. Историческое развитие рассматривается на “макроуровне”, но при этом родоначальником всякой монофилетической группы считается вид. В качестве “наследия” натурфилософии в геккелевой филогенетике присутствует и идея эволюционного прогресса: она отчетливо видна из формы филогенетического дерева, в котором доминирует устремленный ввысь центральный ствол, от которого отходят боковые ветви (рис.2).
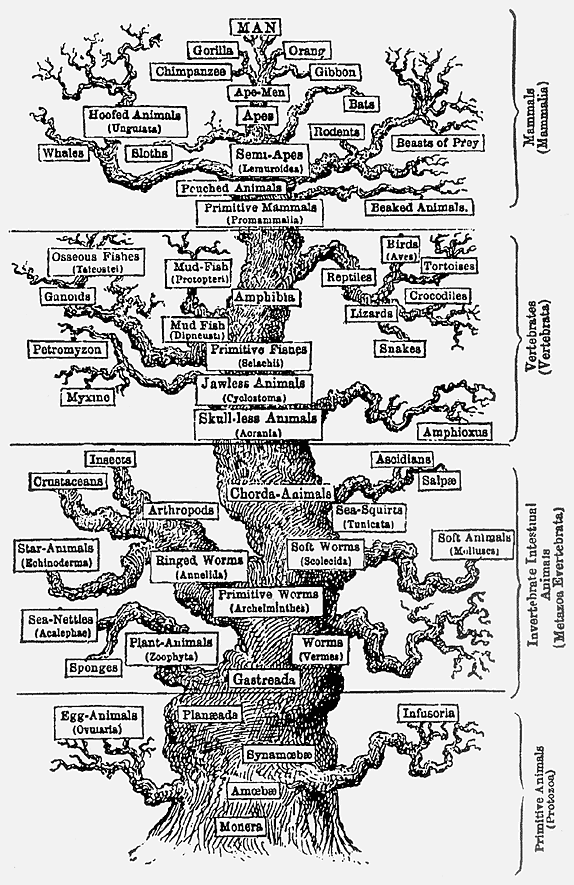
Рис. 2. Одна из ранних версий филогенетического древа по Геккелю.
При изучении семогенетической составляющей филогенеза в основную задачу входит выяснение закономерностей и причин преобразования отдельных структур. В данном случае филогенетическое единство организмов определяется главным образом через общность эволюционных тенденций, которая выражена в единстве направлений и этапности исторического развития. На такое понимание филогенеза во многом повлияли представления об эволюции как однонаправленном процессе, развиваемые некоторыми палеонтологами (Э.Коп, В.О.Ковалевский, Г.Осборн). Этому более соответствует метафора не дерева, а куста или даже травяного поля, на котором каждый “стебель” - филетическая линия - произрастает из своего собственного корня (Л.С.Берг).
Методология классической филогенетики базируется на методе тройного параллелизма, т.е. на максимальном согласовании данных сравнительной анатомии, эмбриологии и палеонтологии. Ключевое значение имеет концепция гомологии, исходно разработанная английским анатомом-платоником Р.Оуэном: подразумевается, что для реконструкции филогенеза важны лишь гомологичные структуры, тогда как аналогичные препятствуют ей. Большое место отводится определению вероятных путей эволюции этих структур на основе критериев, вырабатываемых в рамках перфекционистской (от простого к сложному, от менее к более структурированному) и адаптационистской (от менее к более приспособленным организмам) моделей. Частью классической методологии служит анализ предковых форм: с их помощью разрозненные группы “связываются” в монофилетические образования. Особое внимание уделяется ископаемым материалам, среди которых ищутся реальные предки; при их отсутствии предковые признаки реконструируются.
Важная роль отводится взвешиванию признаков. В классической филогенетике оно всегда трактуется как дифференциальное: разным признакам придается разное значение (разный “вес”). Однако в разных школах филогенетики эта общая концепция трактуется специфическим образом в зависимости от понимания сути филогенеза.
Генеалогическая трактовка предполагает деление признаков на “филогенетические”, не связанные с частными адаптациями, и на “адаптивные”. Подразумевается, что именно первые следует использовать для установления родственных связей между видами, поскольку они унаследованы от предков (ближайших или отдаленных), что свидетельствует о филогенетической общности их потомков.
Адаптационистская трактовка, напротив, придает наибольший вес тем признакам, которые отражают адаптивную специфику данной группы. Фактически эта идея - отголосок прежнего понимания “главной сущности” естественных таксонов. В данном случае существенность морфологических структур связывается с их адаптивной (чаще всего функциональной) значимостью.
В настоящее время традиции классической филогенетики развивает подход, названный филистикой с легкой руки отечественного палеонтолога-энтомолога А.П.Расницына [6]. В ней филогенетическое развитие по-прежнему рассматривается как упорядоченный адаптациогенез, в котором закономерные тенденции преобладают над случайными. Поэтому при характеристике филогенетического единства групп большое значение придается не только единству происхождения, но и единству эволюционных тенденций их представителей как лишнему доказательство монофилии.
Новая филогенетика
Большинство современных реконструкций выполняется в рамках новой филогенетики. Сформировалась она в результате объединения трех подходов, поначалу (в 60-е годы) развивавшихся независимо, - кладистики, нумерической филетики и генофилетики. Ее можно определить как раздел филогенетики, разрабатывающий филогенетические гипотезы на основе кладистической методологии средствами нумерической филетики при использовании молекулярно-генетических данных.
Идеологическое ядро новой филогенетики составляет кладистика, становление которой связывается с именем немецкого энтомолога В.Геннига. Его основополагающая книга была опубликована на немецком в 1950 г., но особую популярность это направление получило после выхода англоязычного издания [7]. Тогда же появился и сам термин “кладистика”.
Методологической основой кладистического подхода служит принцип экономии, который по возможности исключает исходные домыслы об исследуемом явлении. Соответственно, исторические реконструкции проводятся при минимальных априорных допущениях о свойствах эволюционного процесса. Общая концепция эволюции как адаптациогенеза, обязывающая детально прописывать эволюционные сценарии, взвешивать признаки согласно их адаптивной значимости и т.п., оказывается избыточной. Этим обосновывается ключевая для новой филогенетики редукция исторического развития к кладогенезу, в котором нет ни параллельных линий, ни ступеней эволюционного развития.
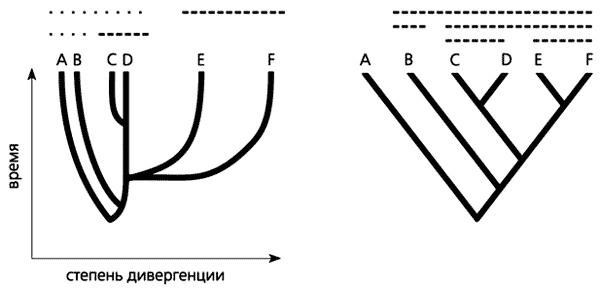
Рис. 3. Стилизованные представления филогенетических схем в современной филогенетике: а - филограмма, б - кладограмма. Цветными линиями показаны парафилетическе группы, штриховыми - голофилетические группы.Такое упрощенное понимание филогенеза существенно изменило его графическое представление. Своего рода символом кладистики, а с ней и всей новой филогенетики стала кладограмма - стилизованное филогенетическое дерево (рис.3). На нем, в отличие от филограммы, показывающей степень дивергенции и уровни продвинутости групп, присутствуют только точки ветвления, отражающие последовательность становления монофилетических групп. Анагенетическая составляющая эволюционного развития отсутствует; поэтому, глядя на кладограмму, нельзя судить об уровнях дивергенции и продвинутости групп организмов.Одна из причин успеха кладистики - эффективная методология филогенетических реконструкций, которая сделала принципиально возможной их воспроизводимость. Ее важная часть - более “узкая” трактовка понятий монофилии и монофилетической группы (см. рис.4). В классической филогенетике монофилетической считается группа, включающая любых потомков данной предковой формы. Очевидно, при выделении на таком филогенетическом дереве монофилетических групп возможны весьма широкие разночтения. В кладистике признаются только голофилетические группы, включающие всех потомков данной предковой формы. Группы, в которых лишь часть таких потомков (парафилетические), не рассматриваются.
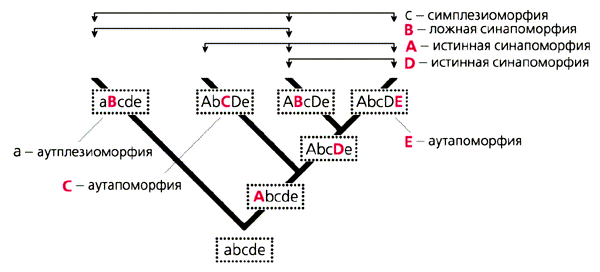
Рис. 4. Категории сходства, связанные с концепцией синапоморфии. Заглавными буквами обозначены апоморфии, строчными - плезиоморфии; цветом выделены апоморфии, определяющие голофилетические группы.Частью этой методологии стал парадоксальный с точки зрения классической филогенетики отказ от учета отношений предок-потомок при обсуждении родства и монофилии. Основанием для этого послужило то соображение, что конкретный предковый вид для подавляющего большинства групп, особенно высокого ранга, нельзя установить. В таком случае более корректна отсылка к сестринской группе, возникшей в результате одного кладистического события вместе с исследуемой. Это дает возможность игнорировать геохронологическую составляющую эволюции и “уравнивать” палеонтологические данные с современными. Такая позиция подводит теоретическую базу под генофилетические реконструкции, в которых предковые формы изначально не нужно исследовать.Основу большинства алгоритмов новой филогенетики составляет принцип синапоморфии, согласно которому монофилию группы можно выявить только через сходство по производному состоянию признака (апоморфии, рис.4). В отличие от этого симплезиоморфия - сходство по исходному состоянию признака (плезиоморфии) - не позволяет определить монофилию группы. При этом особо выделяется синапоморфное сходство, унаследованное от ближайшего предка данной группы (истинная синапоморфия). Напротив, сходство в результате параллельной эволюции (ложная синапоморфия) считается незначимым: это принципиально отличает кладистику от классической филогенетики.
Последнее составляет один из важнейших критериев - значимость признаков: их “вес” обратно пропорционален вероятности параллелизмов и реверсий в эволюции. Чем выше эта вероятность, тем менее значимы признаки, поскольку они с меньшей надежностью позволяют судить о монофилии. Вместо выбора наиболее существенных признаков вводится количественный критерий суммарной оценки значимости сходства: чем большим числом общих апоморфий обладают два таксона, тем выше вероятность их принадлежности к монофилетической группе. Следовательно, чтобы узнать, монофилетичны ли те или иные группировки таксонов, достаточно просуммировать специфические для каждой из них синапоморфии и сравнить результаты. Это позволяет достаточно просто решать конфликты, возникающие между разными признаками, по которым могут получаться разные филогенетические схемы.
Такой критерий стимулировал активное освоение современной филогенетикой количественных методов, которые разрабатывает нумерическая филетика. Сегодня существует своего рода “индустрия” по производству методов количественной оценки сходства организмов и конструирования филогенетических деревьев и их реализации в виде компьютерных программ [8].
В настоящее время наиболее популярны два метода - экономии (парсимонии) и наибольшего правдоподобия. Принципиальные различия между ними состоят в разной трактовке начальных условий реконструкций. Методы первой группы, выделяемые со ссылкой на рассмотренный выше принцип экономии (отсюда их название), предполагают полный отказ от априорного определения эволюции признаков, которые трактуются фенетически. Во второй группе признаки вводятся в качестве элементарных гипотез о семогенезах, в которых определены вероятные направления эволюционных преобразований соответствующих структур. В первом случае итоговая гипотеза оптимальна, если в ней минимально количество параллелизмов; во втором случае - если она наиболее правдоподобно суммирует все эволюционные сценарии для признаков.
Несомненное достоинство нумерической филетики в том, что она позволяет манипулировать большими массивами данных. Морфолог чаще всего способен охватить интуицией лишь ограниченное число структур, которые можно описать несколькими десятками формализованных признаков. Однако без применения количественных техник едва ли можно сравнить десятки таксонов по нуклеотидным последовательностям, состоящим из нескольких тысяч пар оснований. Но здесь кроются и свои “подводные камни”. Так, суммирование нескольких десятков морфологических и нескольких сотен молекулярных признаков приводит к тому, что первые “поглощаются” вторыми. В результате филогенетическая схема почти целиком определяется молекулярно-генетическими данными.
Активная “нумеризация” новой филогенетики все более переносит акцент с теории и методологии реконструкций на их “технологию”. Обсуждаются детали вычисления доверительных интервалов при применении вероятностных критериев, скорость работы компьютерных программ, их ограничения на манипулирование большими массивами данных и т.п. При этом все меньше внимания уделяется биологическому осмыслению результатов, что во многом выхолащивает эволюционный смысл подобных реконструкций.
Особый раздел нумерической филетики составляет компьютерное моделирование филогенетических процессов. Оно позволяет исследовать некоторые свойства исторического развития организмов на основе симуляционных моделей.
Генофилетика (филогеномика) - раздел новой филогенетики, в котором заключения о кладистической истории выводятся на основании анализа исключительно молекулярно-генетических данных. Это новейший этап развития молекулярной филогенетики, связанный с изучением нуклеотидных последовательностей. Теоретические предпосылки для использования таких данных в филогенетике были сформулированы в рамках СТЭ, которая свела эволюцию к изменениям, протекающим на генетическом уровне. На этом основании была выдвинута идея: чем “ближе” признаки к генотипу, тем больше их значение для филогенетических реконструкций. Нечего и говорить, что получение технического доступа к первичной структуре информационных макромолекул было расценено как “молекулярная революция” в филогенетике.
В генофилетике при реконструкции родственных отношений особое значение (со ссылкой на Дарвина) придается изменениям в первичной структуре макромолекул, не связанным с выработкой частных адаптаций. Такая трактовка эволюции вполне согласуется с принципами кладистики, что естественно объединяет последнюю и генофилетику в рамках новой филогенетики. Данное обстоятельство следует подчеркнуть особо: если бы в современной филогенетике доминировала адаптационистская концепция, в ней не нашлось бы места генофилетическим реконструкциям, коль скоро они не интерпретируемы с позиций адаптациогенеза.
В генофилетике организм сводится к первичной структуре биополимеров, каждый элемент которых (нуклеотид) рассматривается как отдельный признак, а их может набираться несколько тысяч. Суждения о родстве базируются на суммарной оценке сходства по этим элементарным признакам, причем чем их больше (чем длиннее последовательность), тем более надежными считаются эти суждения. Поэтому единственный приемлемый метод оценки сходства - количественный.
Для перехода от сходства по молекулярным структурам к родству была разработана концепция молекулярных часов. В ее основу легла так называемая модель “нейтральной эволюции”, предполагающая стохастическую природу накопления нейтральных мутаций в филетических линиях. Ключевым для концепции стало допущение, что ход этих “часов” приблизительно равномерен и одинаков для разных молекулярных структур и в разных группах организмов. Следовательно, если известна скорость накопления мутаций, то можно определять время разделения групп и на этом основании устанавливать между ними генетическое родство.
Количественная оценка различий по молекулярным структурам (генетическая дистанция) дает собственное время дивергенции, напрямую несопоставимое для разных групп организмов. Перевод его в единое для них абсолютное время (в миллионах лет) осуществляется посредством калибровки “молекулярных часов”. Для этого вычисленные дистанции сопоставляются с временем появления в палеонтологической летописи первых представителей монофилетических групп, выявленных молекулярными средствами. Затем проводится перерасчет дистанций в единицах скорости изменения первичной структуры макромолекул и определяется абсолютное время дивергенции.
Отсюда видно, что задача калибровки “молекулярных часов” не решается в рамках самой молекулярной биологии: результат зависит от палеонтологических данных. Здесь очень важны точность определения времени захоронения ископаемых остатков и правильность их отнесения к группам, выделенным по молекулярным маркерам. Кроме того, разные методы оценки генетических дистанций, а также самой калибровки “часов” могут давать разные результаты. Все это служит серьезным основанием для критики исходной упрощенной трактовки концепции “молекулярных часов” [9]. Накопленные данные показывают, что скорость изменения первичной структуры различна для разных макромолекул, а для одних и тех же макромолекул - для разных их участков и в разных группах организмов. Значит, универсальных “молекулярных часов” не существует в силу неравномерности их хода.
Возможности генофилетики существенно ограничены тем, что молекулярно-генетические данные, пригодные для филогенетических исследований, накоплены пока лишь для несколько десятков тысяч из почти 2 млн современных видов, известных науке. А ведь еще есть и многие миллионы вымерших видов, по которым такого рода данные никогда не будут доступны. Все это делает особо актуальной проблему неполноты выборки: чем меньше в исследуемой группе представлено относящихся к ней таксонов, тем менее точна оценка филогенетических отношений между ними.
Приложения новой филогенетики
Многие биологические дисциплины (систематика, биогеография, в последние годы также экология и этология) в той или иной форме используют результаты филогенетических исследований. И в этом смысле их можно считать сферами приложения филогенетики как научной дисциплины.
В настоящее время существует целый класс задач, решение которых так или иначе связано с анализом филогенетического (точнее, кладогенетического) сигнала - информации о происхождении организмов, содержащейся в структуре их сходственных отношений. Оценка этого сигнала равносильна выявлению исторических причин, наряду с другими обусловивших наблюдаемое разнообразие. Очевидно, чтобы выявить названный сигнал, необходимо достаточно полно реконструировать кладистическую историю организмов.
Действительно, всякая развивающаяся система, образно говоря, есть “жертва своей истории”. Это значит, что строение или образ жизни организмов могут быть обусловлены особенностями не только их среды обитания, но и особенностями их предыстории. Например, почему у разных наземных животных разное количество конечностей - четыре у позвоночных, шесть у насекомых, восемь у паукообразных? Ответы вряд ли имеет смысл искать в их условиях обитания. Скорее, они кроются в морфологической организации предковых форм каждой из этих групп, от которых потомки унаследовали свои особенности. Понимание этого привело к формированию исследовательской программы - сравнительной филогенетики. Она изучает проявления исторических причин в особенностях образа жизни, морфологии, поведения и т.п. в разных группах организмов [2, 10].
Чуть ли не с первых шагов своего развития филогенетика тесно связана с исторической биогеографией, предоставляя ей сведения об истории формирования таксонов, составляющих ту или иную локальную биоту. В настоящее время эта связь оформилась в виде кладистической биогеографии, которая целиком опирается на анализ кладистической составляющей филогенеза [11].
Если изначально филогенетика фактически “выросла” из биологической систематики, то сегодня, напротив, новая филогенетика оказывает существенное влияние на систематику, ее принципы и методы. Классическая филогенетика определяет задачи систематики как отражение результатов филогенеза в системе таксонов, характеризуемых филогенетическим единством. Уточнение этого единства дало начало так называемой кладистической систематике, цель которой сводится к “переводу” кладограммы в таксономическую систему.
Парадокс ситуации в том, что это различие, очень незначительное по форме, весьма сильно повлияло на состояние современной систематики. Очень многие таксоны, признанные поколениями систематиков, оказались парафилетическими и потому подлежащими “упразднению”. Это вызвало волну существенных перестроек естественной системы живых организмов и, соответственно, резкую полемику по поводу их правомочности и обоснованности. Типичным примером таких разногласий может служить ситуация, сложившаяся с системой современных представителей рептилий и птиц (рис.5). В классической систематике в качестве естественных групп принимаются линнеевские классы Reptilia (черепахи, гаттерии, лепидозавры и крокодилы) и Aves (птицы). Причина достаточно очевидна: эволюция птиц вывела их за пределы рептилийной адаптивной нормы, так что они утратили единство с ними.
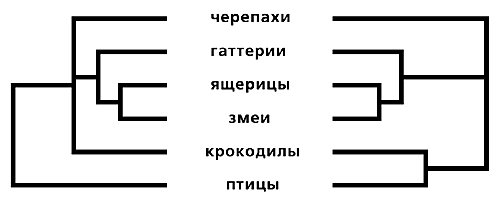
Рис. 5. Классическая (слева) и кладистическая (справа) классификации рептилий и птиц.
Но в генеалогической схеме среди перечисленных групп ближайшие родственники крокодилов - птицы. С точки зрения кладистики, классический таксон Reptilia - парафилетический: в его составе должны остаться только гаттерии и лепидозавры (ящерицы и змеи). Черепах нередко объединяют в класс Parareptilia, а птиц и крокодилов следует выделять в класс Archosauria (куда из ископаемых форм относятся также динозавры и птерозавры).
Генофилетический раздел новой филогенетики разрабатывает так называемую геносистематику, в которой таксоны формируются исключительно на основе генетических данных [12]. Ее заслуга в уникальном фактологическом обеспечении старой идеи “Дерева жизни” - глобальной классификации мира живых организмов. В частности, очень детально прорабатывается таксономическая система для прокариот, приведшая к выделению новых царств живых организмов [13].
Кладистическая систематика не оставила без внимания и таксономическую номенклатуру. В настощее время разрабатывается так называемый “Филокодекс”, идеологическую основу которого составляет принцип монофилии в его “узкой” трактовке [14].
Развитие науки в некотором смысле подобно биологической эволюции. Разнообразие идей, подходов, методов столь же присуще динамично развивающейся научной дисциплине, как разнообразие видов - эволюционирующей биоте. Постоянная конкуренция идей, возникновение и отмирание научных школ, развитие новых направлений - всё это и следствие, и причина научного прогресса. * * *
Филогенетика в этом отношении не составляет исключения. Ее основной вектор развития во все времена задан очевидной “сверхцелью” - все более полной реконструкцией истории формирования биоты. Как достичь ее - вот основная проблема этой дисциплины. Способы ее решения порождают разные подходы к филогенетическим реконструкциям. Формирование новой филогенетики связано с уточнением этой общей задачи и попытками по-новому решить старые проблемы. Но каждый подход действует в рамках своих возможностей, а его неоправданно оптимистичная трактовка часто уводит в неверном направлении. Поэтому было бы ошибочным считать, что предлагаемые сегодня исторические реконструкции - окончательные.
Очевидно, что на смену новой филогенетике придет какой-то другой подход. Можно полагать, что, подчиняясь тенденциям развития эволюционной теории, он будет свободен от избытка редукционизма, присущего современным исследованиям.
Литература
1. Парамонов А.А. Пути и закономерности эволюционного процесса // Современные проблемы эволюционной теории / Ред. В.И.Полянский, Ю.И.Полянский. Л., 1967. С.342-441.
2. Павлинов И.Я. Введение в современную филогенетику. М., 2005.
3. Баранцев Р.Г. Синергетика в современном естествознании. М., 2003.
4. Жерихин В.В. Избранные труды по палеоэкологии и филоценогенетике. М., 2003.
5. Eldredge N., Cracraft J. Phylogenetic patterns and the evolutionary process. N.Y., 1980.
6. Расницын А.П. Процесс эволюции и методология систематики // Тр. Русск. энтомол. об-ва. 2002. Т.73. С.1-108.
7. Hennig W. Phylogenetic systematics. Urbana, 1966.
8. Nei M., Kumar S. Molecular evolution and phylogenetics. Oxford, 2000.
9. Bromham L., Penny D. // Nature Rev. 2003. V.4. №3. P.216-224.
10. Pagel M. // Nature. 1999. V.401. P.877-884.
11. Crisci J.V., Katinas L., Posadas P. Historical biogeography. An introduction. Harvard, 2003.
12. Антонов А.С. // Генетика. 2002. Т.38. №6. С.751-757.
13. Кусакин О.Г., Дроздов А.Л. Филема органического мира. 1. Пролегомены к построению филемы. СПб., 1994.
14. Cantino P.D., Queiroz K. The PhyloCode: a phylogenetic code of biological nomenclature. 2003. URL: http://www.ohiou.edu/phylocode/