 №4, 2002 г.
№4, 2002 г.
 |
© А.Г.Камкин, И.С.Киселева, В.Н.Ярыгин
Фибрилляция, дефибрилляция… А.Г.Камкин, И.С.Киселева, В.Н.Ярыгин Андрей Глебович Камкин, д.м.н., профессор РГМУ.
|
Медицинский термин “фибрилляция”, наверное, помнят все, кто смотрел телевизионный сериал “Скорая помощь”. За констатацией этого неведомого простому смертному состояния пациента следовала команда: “Разряд!” - после чего врачи сообщали о наступлении дефибрилляции, и сердце восстанавливало свою деятельность. Что же такое фибрилляция? Это состояние, при котором клетки сердечной мышцы - кардиомиоциты - сокращаются несогласованно, асинхронно. В результате сердце как насос не выполняет своей функции: кровь не циркулирует по сосудам и кислород не поступает к клеткам организма. Это кислородное голодание особенно опасно для мозга - он живет в таких условиях всего несколько минут.
Смертность людей от различных заболеваний сердца занимает печальное первое место. Одно из последствий сердечных недугов и есть фибрилляция, причины возникновения которой исследуются во всем мире, но все еще так и не установлены. Мы пытаемся понять механизм этого вида аритмий с 1985 г. и, как нам представляется, близки к его раскрытию. О результатах многолетней работы и пойдет речь.
В предыдущей статье мы рассказали о новом типе ионных каналов, открытом примерно 15 лет назад [1]. Теперь мы убедились, что именно эти каналы имеют прямое отношение к фибрилляции.
Механика, электричество и аритмии
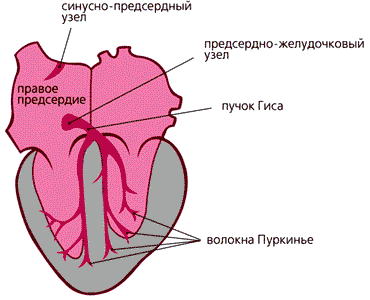
Функция сердца, как известно каждому, - обеспечивать циркуляцию крови по сосудам. Происходит это благодаря последовательному расслаблению предсердий и желудочков (фаза диастолы), а потом их сокращению (фаза систолы). Заканчивается цикл общим расслаблением всех четырех камер. Сокращение сердечной мышцы (миокарда) вызывается электрическим импульсом, который возникает в клетках синусно-предсердного узла, называемого водителем ритма. Далее возбуждение передается по предсердиям и достигает предсердно-желудочкового узла, через пучок Гиса доходит до его конечных разветвлений - волокон Пуркинье - и передается желудочкам. Клетки сердечных камер сокращаются и расслабляются согласованно, и тем обеспечивается отчетливый ритм в деятельности насоса, перегоняющего кровь.
Схема расположения в сердце
водителя ритма и
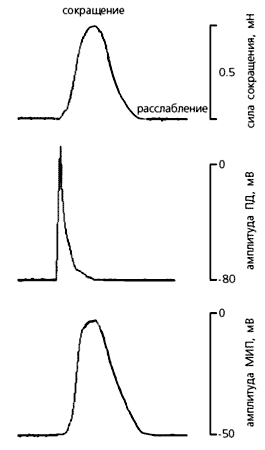
системы, проводящей возбуждение.Невозбужденные клетки миокарда обладают потенциалом покоя (от –70 до –90 мВ), который под действием различных стимулов меняется. Он может стать более отрицательным, тогда мембрана гиперполяризуется, но может и уменьшиться при ее деполяризации. Входящие в клетку катионы (прежде всего Na+ и Ca2+) всегда деполяризуют мембрану, а выходящие (в основном K+) - гиперполяризуют. Быстрый сдвиг потенциала покоя в положительном направлении называют потенциалом действия (ПД). В нем принято выделять три фазы: деполяризации, овершута (перескока) и реполяризации. Во время первой фазы потенциал стремительно нарастает и мембрана теряет свой нормальный заряд; во второй фазе потенциал приобретает положительное значение, а в течение третьей возвращается к исходной величине - потенциалу покоя. Именно потенциал действия и побуждает клетки сердца к сокращению.
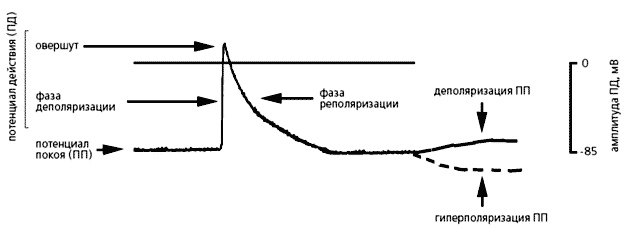
Фазы потенциала действия.
Прямая зависимость сокращения кардиомиоцита от его возбуждения (возникновения ПД), т.е. электромеханическое сопряжение, уже детально изучена. Но существовали клинические данные, свидетельствующие также о наличии в миокарде обратной связи - изменении электрических процессов под действием механических факторов: растяжения миокарда и (или) изменения его сократительной активности. Такая зависимость фактически была установлена еще в 1915 г. английским кардиофизиологом Ф.А.Бейнбриджем. Он показал, что растяжение правого предсердия вызывает учащение ритма сердца у крыс. Соотечественники Бейнбриджа О.Франк и Е.Х.Старлинг, растягивая сердечную мышцу до определенной длины, добились увеличения силы ее сокращения. Из этого можно было бы заключить, что механическая стимуляция ткани сердца должна приводить к изменению мембранного потенциала в клетках. Однако до недавнего времени влияние обратной связи не изучалось и не учитывалось. О ее существовании в 1968 г. лондонский физиолог М.Дж.Лаб высказал лишь предположение, но не доказал его из-за несовершенства экспериментальных методов.
К настоящему времени накопилось множество подтверждающих эту мысль клинических наблюдений. Хорошо известны, например, предсердные аритмии - фибрилляция, пароксизмальная тахикардия, - которые возникают у больных с острым растяжением предсердий или при постепенном увеличении их размеров. Описаны случаи, когда механически индуцированная аритмия возникала у пациентов и без этих патологий, просто при введении катетера в сердце. К различным нарушениям сердечного ритма могут приводить также постоянные механические перегрузки, вызванные артериальной гипертензией, застойной сердечной недостаточностью и хроническим растяжением миокарда. Взаимосвязь механики и электричества подтверждается и давно принятой в медицине механической стимуляцией сердца. Этот непрямой массаж успешно применяют, чтобы восстановить сокращения миокарда или вывести его из состояния фибрилляции.
Таким образом, клинические наблюдения иллюстрируют, что растяжение камер сердца, особенно при его патологии, приводит к аритмиям, но механическая стимуляция может и восстанавливать нормальный сердечный ритм, предотвращать развитие фибрилляции. Поэтому чрезвычайно важно понять, благодаря каким процессам осуществляется механоэлектрическая обратная связь.
Работа кардиомиоцитов
В физиологических условиях во время расслабления сердечных камер (т.е. в диастоле) механическое воздействие на миокард приводит к изменению потенциала покоя на мембране кардиомиоцитов и следовательно протекающего через нее тока. Этот ответ обусловлен работой механочувствительных ионных каналов (в научной литературе их называют механосенситивными и обозначают аббревиатурой МСК, которую мы сохраним в данном тексте).
Зависимость электрической активности кардиомиоцитов от механических воздействий на них мы начали изучать давно. В экспериментах имитировали нормальные сокращения этих клеток в зависимости от величины растяжения ткани, для чего растягивали фрагменты ткани миокарда с силой, равной физиологической, а затем превышали эту силу и измеряли мембранный потенциал. В результате выяснилось, что дополнительное растяжение (1.5 мН) ткани правого предсердия приводит к изменению обычной формы потенциала действия: в фазе реполяризации, на уровне 90%, начиналась деполяризация мембраны. Если растяжение продолжали, возникал внеочередной потенциал действия. В опытах на ткани предсердий, взятой от животного, перенесшего инфаркт левого желудочка, достаточно было превысить силу растяжения над физиологическим значением всего на 0.1 мН, чтобы вызвать более серьезные изменения: увеличивалась длительность реполяризации уже на двух уровнях (50 и 90%); вдвое большая сила приводила к дополнительному потенциалу действия; если же она достигала 0.3 мН, начиналась фибрилляция предсердия [2]. Сходные результаты дали эксперименты на фрагментах желудочков сердца [3].
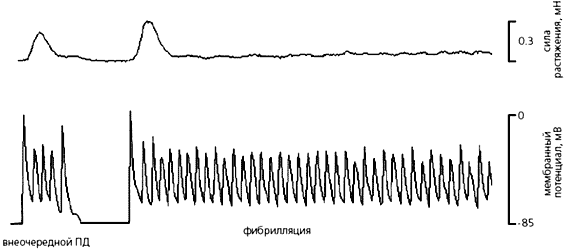
Регистрация механической (мН) и биоэлектрической (мВ) активности ткани правого предсердия здоровой крысы во время механического воздействия на препарат. Растяжение, превышающее физиологическую величину (принята за ноль на механограмме, вверху), вызывает обычный потенциал действия, и он остается почти неизменным, пока приложенная сила не достигнет 2 мН. Растяжение такой силы вызывает отчетливый внеочередной потенциал действия и сокращение ткани. Когда дополнительная растягивающая сила снимается, амплитуда потенциала возвращается к норме. Стрелками указаны моменты приложения растягивающей силы.Итак, при растяжении миокарда или изменении силы сокращений сначала наступает деполяризация мембраны и меняется длительность потенциала действия главным образом за счет аномально протекающей реполяризации (на разных ее уровнях). Затем на этой же фазе может начаться повторная деполяризация, в результате чего нарушается нормальный ритм сердца - возникают аритмии, вплоть до фибрилляции [4].
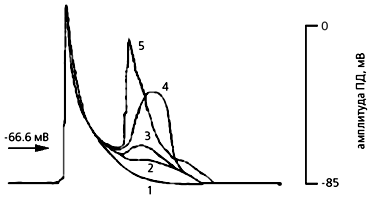
Зависимость величины мембранного потенциала от степени растяжения препарата предсердной ткани крысы, перенесшей инфаркт миокарда. Кривая 1 соответствует потенциалу при физиологическом растяжении, на каждом следующем шаге (кривые 2-5) сила увеличена на 0.05 мН. Когда растяжение приводит к критическому уровню деполяризации (равному –66.6 мВ), возникает внеочередной потенциал.
Запись механической (мН) и биоэлектрической (мВ) активности кардиомиоцитов из предсердия крысы, перенесшей инфаркт, при растяжении клеток. Когда растягивающая сила увеличена всего на 0.2 мН (вверху), появляются внеочередные потенциалы действия (внизу). Дальнейшее растяжение с силой, превышающей 0.2 мН, приводит к фибрилляции.Поскольку эффект растяжения ткани миокарда блокировался ионами гадолиния - ингибиторами механочувствительных каналов, - напрашивался вывод об их участии в описанных событиях [5]. Но чтобы доказать это, необходимо было исследовать ионные токи и сами МСК не на ткани сердца, а на отдельных жизнеспособных клетках. Для растяжения единичного кардиомиоцита и измерения в нем тока, мы разработали специальный прием, который и использовали в дальнейших экспериментах на изолированных клетках из желудочков сердца [4].
Результаты оказались сходными с теми, что были получены на ткани предсердий: чем с большей силой растягивалась клетка, тем значительней для нее были последствия; кардиомиоциты больного сердца оказались чувствительнее к растяжению, чем здорового, независимо от того, были это клетки морской свинки или человека; сильнее, по сравнению с кардиомиоцитами здоровой крысы, отвечали на растяжение клетки животного, страдающего артериальной гипертензией. Сила ответа зависела также и от возраста: для изменений величины потенциала покоя и длительности потенциала действия требовалось гораздо большее растяжение клеток сердца молодых крыс, чем старых.
Таким образом, прямое растяжение как предсердных, так и желудочковых кардиомиоцитов вызывает деполяризацию их мембраны и увеличение фазы реполяризации на уровне 90%. Если оба эти процесса достигают критических величин, возникает дополнительный потенциал действия, что приводит к экстрасистоле - внеочередному сокращению сердечных камер.
Суммарный ток, протекающий через мембрану кардиомиоцита, тоже увеличивался по мере растяжения клетки и исчезал при возвращении ее длины к исходной. Какими ионами вызван ток, мы установили в экспериментах, заменяя один ион на другой, например K+ на Cs+, Na+ на более крупный катион триэтиламмония, а Cl– на F–. В результате выяснилось, что при растяжении клетки за ток ответственны ионы натрия, входящие в нее через механочувствительные каналы. В опытах по исследованию токов изолированных кардиомиоцитов использовались образцы, взятые из тех же объектов, т.е. здоровых морских свинок и крыс (молодых и старых), людей с заболеваниями сердца и крыс с гипертензией. И здесь картина была такой же, как в экспериментах по изучению потенциала действия: чувствительность клеток к экспериментальному растяжению увеличивалась с возрастом животных, но еще сильнее была связана с гипертензией. Такое повышение чувствительности мы объясняем увеличением на мембране числа механочувствительных каналов [6].
Итак, полученные нами результаты позволяют заключить, что не только электрическое возбуждение приводит к сокращению или расслаблению клеток сердца, т.е. к их механическому изменению, но и наоборот - на механическое воздействие кардиомиоциты отвечают электрической активностью. Однако если прямая связь обеспечивает нормальный сердечный ритм, то обратная сказывается противоположным образом - нарушением ритмики при всех патологиях.
Считается, что механические раздражения воспринимаются лишь кардиомиоцитами - электровозбудимыми сократительными клетками. Мы же доказали, что в обратной связи, особенно при заболеваниях сердца, могут участвовать не только они, а еще и фибробласты - немышечные электроневозбудимые клетки.
Новая роль фибробластов
В нормальном миокарде группа немышечных клеток состоит из разных типов, но преобладают в ней фибробласты, которым приписывается роль опорных структур. Однако, судя по накопившимся сведениям о других сторонах метаболической активности фибробластов, они участвуют и в регуляции работы сердца [4, 7]. К такому предположению мы пришли еще в 1986 г. Изучение электрофизиологических характеристик фибробластов бьющегося сердца и их межклеточного взаимодействия привело нас к мысли, что деятельность миокарда обеспечивается не одними лишь кардиомиоцитами, а их совместным функционированием с этими опорными клетками. В таком случае работа и здорового сердца, и измененного болезнью должна выглядеть иначе, чем принято считать.
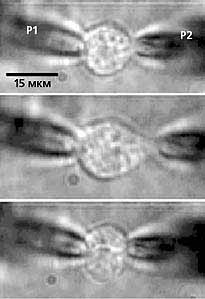
В сердце млекопитающих не так много фибробластов, примерно 5-10%, но в зоне синусного узла их количество достигает 45-75%. Эти немышечные клетки отличаются от кардиомиоцитов целым рядом электрофизиологических особенностей. К их числу относятся низкая величина потенциала покоя (в среднем –25 мВ) и высокое входное сопротивление - около 1 ГОм. В ритме спонтанных или вызванных сокращений миокарда фибробласты генерируют механоиндуцированные потенциалы (МИП), форма которых совершенно не похожа на форму потенциала действия кардиомиоцитов. Амплитуда МИП фибробластов либо соизмерима с величиной потенциала покоя кардиомиоцитов, либо значительно меньше его. Характерно, что МИП никогда не перескакивает через нулевое значение, т.е. в его форме отсутствует овершут. В экспериментах величина потенциала реверсии зависит от степени растяжения препарата, но при стандартной силе воздействия она составляет всего –5 мВ. В одинаковых экспериментальных условиях механоиндуцированный потенциал фибробластов генерируется позднее, чем потенциал действия кардиомиоцитов: в сердце лягушки на 90 мс, крысы - на 10, человека - на 40 мс. По длительности МИП совпадает со временем сокращения препарата.
Справа - Механограмма (вверху) и потенциалы, возникающие в кардиомиоците (в середине) и фибробласте (внизу) правого предсердия крысы. Видно, что потенциал действия (ПД) кардиомиоцита приводит к сокращению ткани сердца, а механоиндуцированный потенциал (МИП) фибробласта появляется при сжатии этой клетки во время сокращения ткани.
В нормальных физиологических условиях МИП фибробластов возникает в ответ на сокращения сердца. В период систолы фаза нарастания этого потенциала связана с активацией неселективных механочувствительных каналов, через которые в клетку поступают различные ионы, в том числе Ca2+. Если увеличивается концентрация ионов кальция, входящий ток резко сдвигает потенциал покоя фибробластов и возникает МИП. Его амплитуда растет до той поры, пока не начнется расслабление миокарда, ведущее к инактивации этих каналов. Нисходящую фазу МИП обусловливает, видимо, ток, который возникает за счет выхода из клетки K+ через калиевые каналы, активируемые кальцием. Но возможно, эта фаза связана с инактивацией МСК.
Фибробласты специфически, иначе, чем кардиомиоциты, реагируют и на искусственную поляризацию мембраны, и на растяжение ткани сердца. Так, вызванная гиперполяризация внутренней мембраны фибробластов спонтанно сокращающегося сердечного фрагмента обычно приводит к увеличению амплитуды МИП, а деполяризация - к уменьшению. Примечательно, что первый сдвиг зависит от величины, на которую сместился потенциал покоя фибробластов, а второй - от степени искусственной поляризации.
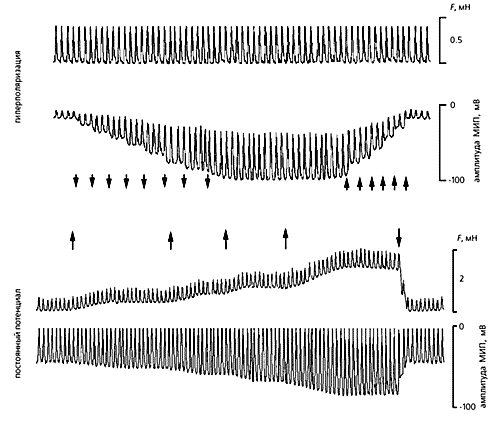
Дополнительное растяжение ткани спонтанно сокращающегося фрагмента правого предсердия вызывает сдвиг потенциала покоя в сторону гиперполяризации мембраны фибробласта. Соответственно увеличивается и амплитуда МИП, причем ее величина прямо связана со степенью гиперполяризации. Если растяжение устранить, потенциал покоя возвратится к исходной величине. Столь выраженная реакция фибробластов на растяжение свидетельствует, на наш взгляд, об их участии в осуществлении механоэлектрической обратной связи.
Реакции фибробластов из предсердия крысы на искусственную поляризацию мембраны и растяжение клетки. Растянутая ткань ритмично сокращается (верхняя механограмма) с силой около 0.4 мН. Амплитуда потенциала (МИП), индуцированного таким воздействием, увеличивается на каждом шаге искусственного усиления гиперполяризации мембраны (моменты усиления отмечены стрелками, направленными вниз) и уменьшается, когда гиперполяризация так же ступенчато снимается (стрелки, направленные вверх). Если же потенциал мембраны поддерживать на постоянной величине, а силу растяжения увеличивать несколько раз, то по мере роста растягивающей силы амплитуда МИП тоже увеличивается. Все это подтверждает участие фибробластов в механических и электрических событиях в сердце.Роли стромы (соединительной ткани, в состав которой входят фибробласты), в нормальном и больном сердце уделялось мало внимания. А между тем еще в 1971 г. описаны случаи ишемии и инфаркта миокарда, сопровождавшиеся воспалительным процессом ткани, который привел к частичному или полному повреждению кардиомиоцитов, но работа фибробластов активировалась. Эти клетки превратились в миофибробласты с более крупным ядром, обильным эндоплазматическим ретикулумом и актиновыми филаментами. Наличие последних структур вполне допускает мысль, что такие трансформированные фибробласты способны сокращаться.
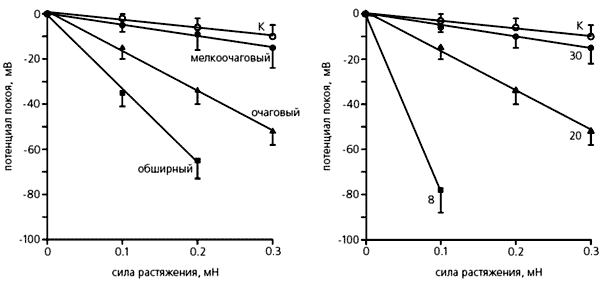
Мы изучали электрические характеристики фибробластов правого предсердия крысы после инфаркта (его вызывали перевязыванием коронарной артерии) миокарда и установили, что потенциал покоя и сопротивление мембраны зависели от размера зоны, охваченной инфарктом. Чем больше она была, тем больше сдвигался потенциал покоя фибробластов: от –22 мВ у контрольных животных и –25.8 у крыс с мелкоочаговым инфарктом до –46.5 мВ - с обширным.
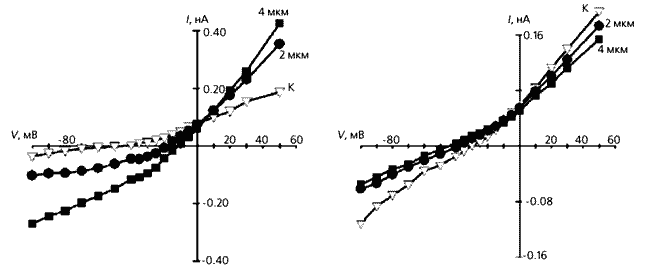
Изменение потенциала покоя фибробластов при растяжении ткани правого предсердия крысы с инфарктом миокарда левого желудочка. Слева: изменения, зависящие от размера инфарктного очага, справа - от количества дней, прошедших после инфаркта; К - контрольные измерения, проведенные на фибробластах здоровых крыс. Видно, что потенциал покоя снижается по мере увеличения очага болезни, т.е. эти “опорные” клетки становятся более чувствительными к механическому воздействию. Отчетливо заметна и повышенная чувствительность фибробластов к растяжению ткани в ранние сроки после инфаркта.Как могут сказаться более отрицательный потенциал покоя и повышенная амплитуда МИП фибробластов? По нашему мнению, первый эффект способен снизить спонтанную активность клеток водителя ритма, а второй - вызвать удлинение потенциала действия соседних кардиомиоцитов.
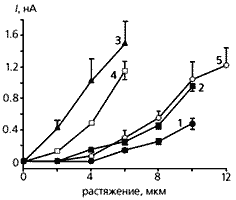
У животных с разной величиной инфарктной зоны отличалась также и чувствительность фибробластов к механическому фактору. Примечательно, что проявлялась та же зависимость, которую мы установили в опытах по измерению электрических характеристик. Однако реакция фибробластов была связана и со временем, прошедшим после инфаркта - чем меньшее количество дней отделяло наступление болезни от начала опыта, тем сильнее сдвигался потенциал покоя фибробластов даже при их незначительном растяжении.
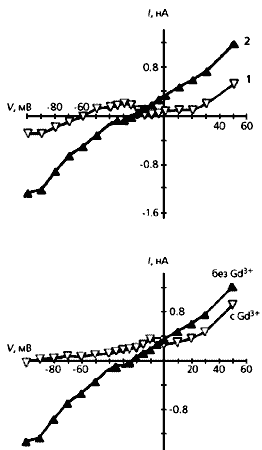
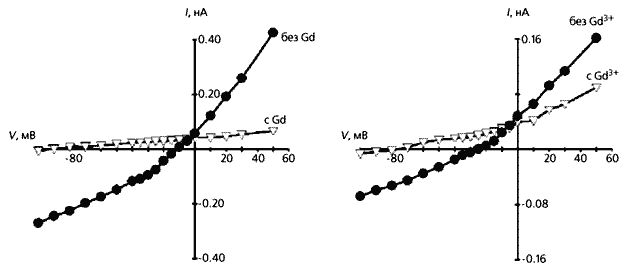
Реакция изолированного фибробласта на сдавливание (слева) и растяжение. Видно, что в первом случае электрическая проводимость увеличивается по сравнению с нормальной клеткой (К), и чем больше она сжата, тем сильнее ток, но потенциал покоя при этом уменьшается. Ответ фибробласта на растяжение противоположен - его мембранная проводимость снижается, а потенциал покоя увеличивается.Мы объясняем повышенную чувствительность фенотипически измененных инфарктом фибробластов (т.е.миофибробластов) к механическому воздействию увеличением числа МСК в мембране (их участие подтверждается действием ингибитора каналов - ионами гадолиния). Даже небольшое растяжение способно активировать дополнительное количество МСК и тем повысить входящий ток. А поскольку возрастает и концентрация свободного внутриклеточного кальция, открываются зависимые от этого иона калиевые каналы и мембранный потенциал сильно сдвигается в сторону гиперполяризации.
Влияние ионов гадолиния на сжатый (слева) и растянутый фибробласт. Ингибирование тока этими ионами проявляется на той и другой клетке и, кроме того, приводит к увеличению потенциала покоя.Как же передаются потенциалы между фибробластами и кардиомиоцитами? В сердце фибробласты взаимодействуют между собой преимущественно через высокопроницаемые контактные зоны мембран, содержащие кластеры специфических межклеточных каналов. Они резко снижают сопротивление мембраны соседствующих клеток и делают возможным их электрическое взаимодействие. Такие же каналы, но распределенные диффузно, существуют и в зоне контакта фибробластов с кардиомиоцитами. Изменение, скажем, мембранного потенциала первых клеток может вызвать такой же отклик в соседних с ними кардиомиоцитах. Вокруг клеток водителя ритма в сердце образованы кольца из фибробластов. Именно их мембранный потенциал увеличивается по абсолютной величине и сдвигает потенциал покоя кардиомиацитов. В результате такого смещения у клеток водителя ритма уменьшается частота возникновения потенциала действия, а значит, и частота сердечных сокращений. Как раз это и наблюдается после инфаркта.
Чтобы понять, как фибробласты отзываются на механический стимул (растяжение или сдавливание), мы измеряли токи, протекающие через мембрану изолированных, единичных клеток [8]. По результатам выяснилось, что сдавливание всегда приводит к смещению потенциала покоя в сторону деполяризации и реверсии входящего тока на выходящий. Четко проявлялось и ингибирующее действие гадолиния. Далее следовало узнать, потоком катионов или анионов обусловлены выявленные эффекты. Для этого мы заменили ионы K+ на ионы Cs+ в условиях сдавливания клетки и обнаружили ту же тенденцию, что и в предыдущих экспериментах: потенциал покоя смещался в сторону деполяризации (при сжатии на 4 мкм он сдвигался от –32 мВ в среднем до –13 мВ), полярность дифференциальных токов тоже менялась. В экспериментах по выявлению роли анионов Cl– мы не обнаружили их влияния ни на потенциал мембраны, ни на токи. Следовательно, ответ фибробласта на сдавливание осуществляется через катионные потоки.
А что происходит при растяжении этой клетки? Оказалось, что оно приводит к гиперполяризации мембраны и значительно уменьшает силу тока по сравнению с контролем. (Интересно, что ионы гадолиния действуют еще сильнее, чем механический стимул.) Влияние анионов хлора здесь тоже не было обнаружено.
Таким образом, результаты, полученные на изолированных фибробластах и на многоклеточных препаратах, совпадают. Этим мы доказали свое исходное предположение о том, что фибробласты сердца отвечают на механическую стимуляцию изменением ионных токов. Обусловлено это, как мы полагаем, неселективной катионной проводимостью. Поскольку механочувствительные каналы фибробластов реагируют на направление приложенной силы (инактивируются растяжением и активируются сдавливанием), можно полагать, что сокращающиеся в систоле кардиомиоциты действуют на лежащие между ними фибробласты как биологическое сдавливающее устройство.
Итак, кардиомиоциты и в еще большей степени фибробласты эффективно преобразуют механическое раздражение в электрические ответы, причем работа первых клеток модулируется вторыми. В здоровом сердце растяжение кардиомиоцитов, приводящее к деполяризации их мембраны, и растяжение фибробластов, вызывающее гиперполяризацию, находятся в равновесии. При патологии же реакция на такое механическое раздражение выражена особенно сильно - но по-разному - в тех и других клетках. Если величина гиперполяризации фибробластов больше, чем деполяризации кардиомиоцитов, сердечный ритм становится реже и даже может прекратиться. И наоборот, если преобладает последняя, начинается аритмия и может развиться фибрилляция. Математическое моделирование полностью подтверждает такие выводы из экспериментальных данных.
Выявленное нами взаимодействие этих двух типов клеток и его влияние на работу сердца важно не только для фундаментальной науки, но и для практической медицины. Если учесть, что медленное хроническое растяжение миокарда за счет внутрисердечного давления вызывает сдвиги потенциалов и как результат - нарушение ритма, то можно понять, что даже изменение позы больного, перенесшего инфаркт, может привести к драматическим последствиям. Мы близки к тому, чтобы перенести исследования в клинику, но до этого предстоит изучить электрофизиологические характеристики кардиомиоцитов и фибробластов человеческих сердец - здоровых и с разными патологическими изменениями. Исходя из наших представлений об участии в механоэлектрической обратной связи специфических ионных каналов, необходимо найти для них селективный блокатор, который прерывал бы эту связь и тем восстанавливал нормальную работу сердца. Пока такого блокатора не существует. Но им могли бы стать антитела к механочувствительным каналам, правда, сначала их нужно “клонировать”. Можно блокировать одно из звеньев той цепочки, которая приводит к экспрессии МСК, однако эту цепочку прежде необходимо выявить. Поиском блокаторов мы уже занимаемся, и когда эксперименты принесут желаемый результат, не понадобятся ни электрический разряд, ни прямой механический массаж, чтобы вывести пациента из состояния фибрилляции. Она просто не возникнет.
Литература
1. Камкин А.Г., Киселева И.С., Ярыгин В.Н. // Природа. 2002. №3. С.13-20.
2. Kamkin A., Kiseleva I., Wagner K.D. et al. // J. Mol. Cell. Cardiol. 2000. V.32. P.465-477.
3. Kiseleva I., Kamkin A., Wagner K.D.et al. // Cardiovasc. Res. 2000. V.45. P.370-378.
4. Камкин А.Г., Киселева И.С. // Успехи физиол. наук. 2000. Т.31. С.51-78.
5. Kamkin A., Kiseleva I., Isenberg G. // Cardiovasc. Res. 2000. V.48. P.409-420.
6. Камкин А.Г., Киселева И.С., Ярыгин В.Н. // Успехи физиол. наук. 2001. Т.32. С.75-104.
7. Камкин А.Г., Киселева И.С. // Успехи физиол. наук. 1998. Т.29. С.72-102.
8. Kamkin A., Kiseleva I., Husse B., Isenberg G. // Europ. J. Physiol. 2001. V.441. P.15-4, R191.

Март 2002