№3, 2006 г.
© Берман Д.И.
Как зимуют муравьи на Колыме?
Д.И. Берман
Даниил Иосифович Берман, к.б.н., зав. лаб. биоценологии Ин-та биологических проблем Севера ДВО РАН.Россия, как известно, самая большая заснеженная страна мира. Доля ее площади с положительными температурами зимой ничтожна, а вечная мерзлота занимает почти 60%. На всем этом пространстве каждый год зимуют в оцепенении мириады беспозвоночных животных тысяч разных видов. И только для немногих из них выяснено, как они переживают зиму, как защищаются от холода [1, 2]. Исследователей же холодоустойчивости животных ныне в России - по пальцам одной руки перечесть… Масштабу явления явно не соответствует степень изученности, прикладываемые усилия и оказываемое ему внимание.
Наши предшественники установили, что два вида изученных ими муравьев не выносят замерзания [3]. Казалось, что это свойство всех муравьев. Так как же они зимуют вблизи полюса холода Северного полушария? Пытаясь ответить на этот вопрос, мы много зим и лет работали с муравьями в 500 км от Магадана в верховьях Колымы на нами же построенном стационаре “Абориген”. Осмысление - дело не быстрое, сейчас оно, похоже, пришло, и предлагаемая статья - попытка поделиться им.
В основе подхода лежала триада:
- экология зимовки муравьев (биотопическое распределение, устройство гнезд, зимующие онтогенетические стадии и т.п.),Такой круг задач в одиночку не решается. Над тем трудилась вся наша группа - мирмеколог З.А.Жигульская; физиолог, а в прошлом ядерный физик А.Н.Лейрих, микроклиматолог А.В.Алфимов, начальник стационара В.А.Бельгер, его жена и наша сестра-хозяйка А.П.Бельгер, рабочий, ныне инженер А.А.Поплоухин, водитель А.А.Лейман и др. На последних четверых лежало все, чего остальные просто не умеют: добыть зимой электричество, тепло, воду - словом, все жизнеобеспечение. Когда “за бортом” ниже –50°С, ни закон Кулона, ни сообщающихся сосудов, как выяснилось, на Колыме почему-то не работают. А у этих в ту пору молодых работали. Главное же было в понимании, что лаборатории существуют не только для того, чтобы по одной ведомости зарплату получать…- микроклимат и температурные условия зимовки каждого вида,
- устойчивость муравьев к холоду (температуры максимального переохлаждения, пороги переносимых температур, биохимия криопротекторов и т.п.).
Муравьи как модельный объект
На северо-востоке Азии, как и везде на Севере, видов беспозвоночных много меньше, чем на Юге, зато холод делает картину максимально рельефной. Где, как не здесь или в Антарктиде, заниматься экологией зимующих беспозвоночных? В Антарктиде, безусловно, хуже - там нет муравьев.
Муравьи - чуть ли не идеальный модельный объект для подобных исследований. Как и многие другие беспозвоночные, они разнообразны, населяют практически все ландшафты Северо-Востока, кроме тундр и высокогорий. Но в отличие от всех прочих, муравьев для зимних экспериментов не надо собирать осенью по штуке и укладывать на зимовку в наилучшие (с точки зрения экспериментатора) условия в лаборатории. Не трогая животных осенью, мы даем им возможность пройти естественный режим весьма сложной подготовки к зимовке. Достаточно пометить муравейники длинными вешками (чтоб были видны над снегом), в нужное время зимой выдолбить их целиком из грунта, перенести в …нет-нет, не в теплую лабораторию, а в холодную комнату, чтоб предотвратить тепловой шок, не дать ожить раньше, чем требуется программой эксперимента, не сдвинуть весьма лабильную биохимию криопротекторов и т.д. В таком помещении с приоткрытыми окнами и закрытыми кошмой батареями, чтоб не грели, поддерживались компромиссные –10…–15°С - муравьям не опасно, исследователю терпимо. Монолит с муравейником осторожно вскрывался, муравьи извлекались, раскладывались по бюксам небольшими порциями, используемыми для экспериментов по мере надобности.
Характерный пейзаж бассейна верховий Колымы. На всем пространстве панорамы, кроме горных тундр, обитают муравьи гипоарктического комплекса (C.herculeanus, F.gagatoides, L.acervorum). На заднем плане по центру - среди лиственничников южного склона на полянах и рединах поселяются F.lemani. На днище долины (за пределами поймы) обычна F.exsecta, реже - F.sanguinea. На шлейфах склонов всех экспозиций в моховых займищах можно найти M.kamtschatica. В пойме реки по галечникам и пескам многочисленны поселения F.picea и M.bicolor.“Чтобы сделать рагу из зайца, надо, как минимум, иметь кошку”. Чтобы работать с муравьями зимой, надо выдолбить из грунта муравейник. Полагаю, кошку добыть проще… Представьте, что такое выколоть монолит объемом в четверть куба при колымских морозах, да быстро, чтоб не замерз (разумеется, муравейник), да еще под покрикивание “надсмотрщика”, заботящегося о сохранности содержимого кома с муравейником больше, чем об отваливающихся от холода ногах и душевном равновесии нецензурно капризничающих работающих. Потом надо погрузить на нарты, потом затащить в “холодную”. В одиночку всего этого не сделать, и в роли добытчиков выступало все мужское население лаборатории. Описываю столь подробно лишь ради того, чтобы стало понятно: криоэкология на Колыме - наука, сопряженная в первую очередь с кайлом, лопатой и свирепыми морозами, а учитывая место действия, еще и добровольная каторга. Лаборатория, измерения температуры насекомых тончайшими термопарами, самописцы, низкотемпературные холодильники, биохимические анализы и прочие атрибуты - все это уже потом, всласть.Здесь и далее фото автора
Местообитания, устройство гнезд и зимовка
От верховьев Колымы, где расположен стационар “Абориген”, рукой подать до полюса холода Северного полушария (метеостанции “Делянкир” на междуречье Колымы и Индигирки, “Оймякон” и “Мома” на Индигирке) с его нормальными для декабря-февраля –60°С, а то и более низкими температурами. На Колыме не намного лучше: –50°С обычны (при такой температуре работать на открытом воздухе запрещено), –35°С почитают за оттепель.
Снега мало, поэтому и под ним холодно. Среднегодовая температура воздуха на всем востоке страны на юг до бассейна Амура отрицательна, значит - повсеместна мерзлота, прорезанная узкими полосами таликов вдоль рек. Температура мерзлоты, измеряемая на глубине, где не сказываются сезонные колебания (3-4 м), невелика, обычно всего-то –4…–6°С. Правда, никто там и не живет. В тощей же почве, мощность ее редко превышает 15-20 см, температуры на глубине 10 см в ноябре при бесснежье падают до –20…–30°С. По мере роста снежного покрова в почве теплеет, но ненамного, ибо 30-50 см снега принципиально не меняют дела. Самое теплое место из найденных нами - в горных тундрах (выше 1200 м над ур.м.) под двухметровыми надувами снега. Тут зарегистрированы аж –9°С; только под надувами тоже мало кто живет (разве что полевки, весной выселяющиеся из-под тающих снежников в летние норы) - и без того короткий теплый сезон здесь непозволительно урезан.
Лето тоже не балует, оно сокращено до двух месяцев - до середины июня еще весна, а к 20 августа следы осени искать не надо, они везде. Выше границы леса (800-1000 м) к этому времени непременно ложится снег.
Вечная мерзлота может препятствовать устройству камер муравьев на глубине, достаточной по температурным условиям для зимовки. Конечно, речь идет не о “сухой мерзлоте” на хорошо дренируемых склонах, проявляющейся только отрицательными температурами на глубине 2.5-3.5 м да отдельными кристалликами льда, не способными скрепить грунт в ледовый бетон. Она муравьям не помеха.
Другое дело - слитная, почти каменной твердости мерзлота, мешающая муравьям жить, прежде всего, в теплое время года. Над ней температуры близки к нулю и, как на водоупоре, всегда сочится вода. Наиболее ярко мерзлота выражена на северных склонах, заболоченных шлейфах склонов всех экспозиций и днищах долин. Сезонное оттаивание на северных склонах местами не превышает 30-40 см. Забегая вперед, отметим, что все виды рода Formica, кроме полярного муравья (F.gagatoides), как наименее холодостойкие и строящие гнезда не вблизи поверхности почвы, в биотопах с неглубоко оттаивающей водоупорной мерзлотой обычно не живут. Поселения же F.exsecta могут все же встречаться, но лишь в местах с особо благоприятным сочетанием условий. А на биотопическом распределении мезофильных видов, располагающих гнезда вблизи поверхности почвы, даже водоупорная мерзлота никак не сказывается. Будто бы ее и нет. Свидетельство тому - очень высокая численность гнезд некоторых видов.
Лидеры
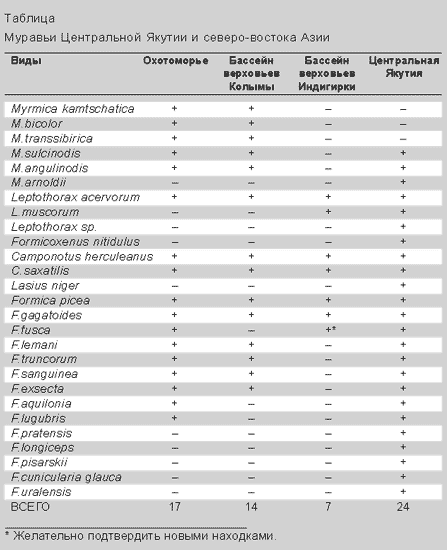
После приведенного описания не должно казаться странным, что муравьи, весьма разнообразные на юго-востоке нашей страны (в Приморье и Приамурье их примерно 80 видов [4]), на северо-востоке Азии, включая бассейн Индигирки, представлены лишь 18 широко распространенными, в большинстве своем транспалеарктическими видами (таблица). Обеднение почти в 4.5 раза! Примерно половина из них обычна. Таким образом, континентальный горно-лесной северо-восток Азии и в отношении муравьев подтверждает репутацию региона с жестким климатом.
В числе лидеров - лишь три вида. Они повсеместны, в том числе в зональных - холодных и сырых биотопах: в моховых лиственничных редколесьях и зарослях кедрового стланика, на болотах и т.д. Это крошечный (3 мм) Leptothorax acervorum и самый крупный (свыше 10 мм) из палеарктических муравьев - древоточец Camponotus herculeanus, а также мелкий (5-6 мм) среди муравьев рода Formica - полярный муравей. Названные муравьи - такие же атрибуты Гипоарктики, как шикша, багульники, карликовые березки и др.
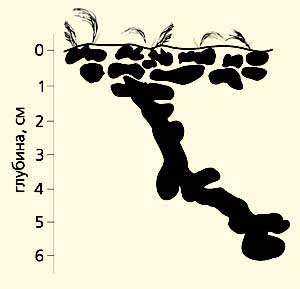
Гнездо Leptothorax acervorum. Поверхностный слой почвы, а с ним и гнезда муравьев подвержены всем превратностям погоды и зимой, и летом. Однако муравьи этого вида, как и L.muscorum, без ущерба переносят охлаждение вплоть до –40°С, а личинки - еще и ниже. Они прекрасно адаптированы и к летним, весьма скудным по теплообеспеченности условиям (кроме как на степных участках), и к зимним холодам. Гнезда L. acervorum имеют примитивнейшее строение; многочисленные (до 10 на 1 м2) зимовочные камеры находятся на глубине всего лишь 5-7 см. Древоточец также прокладывает ходы неглубоко (в 5-15 см от поверхности почвы), причем несмотря на условия, в том числе и уровень зеркала мерзлоты. Гнезда обычно строит в мертвой, сухой древесине; на протяжении 0.5-1.5 м от комля непременно на нижней поверхности корней под корой или в толще древесины может находиться до двух-трех десятков камер, которые и служат зимовочными. Именно такое их расположение - свидетельство безразличия обоих видов к низким температурам. Здесь они опускаются до –25…–28°С, а –20°С держится в гнездах около двух месяцев. Полярный муравей устраивает зимовочные камеры чуть глубже - в 10-15 см, редко 25 см от поверхности почвы, где, разумеется, помягче: минимумы порядка –18…–22°С, но –15°С регистрируется в течение месяца.

Зимовочные камеры муравья-древоточца иногда расположены и в тонких корнях диаметром не более 10 см.
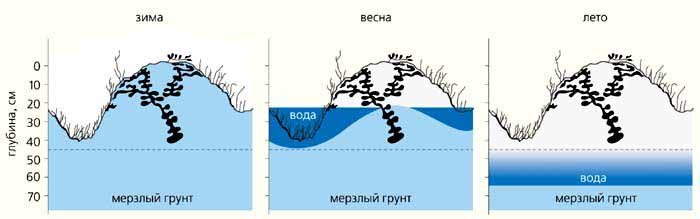
Муравей Myrmica kamtschatica поселяется исключительно в сфагновых и кустарничково-сфагновых редколесьях на шлейфах склонов. Там зеркало мерзлоты даже в период максимального оттаивания опускается не глубже 60-70 см. Соответственно и гнезда устраивает вблизи поверхности моховых подушек. Холодоустойчивость позволяет ему, как и полярному муравью, жить повсеместно. Однако в отличие от него камчатский муравей капризен и за пределами названных биотопов не встречается.
Гнезда Myrmica kamtschatica в разные сезоны. Только к июлю мерзлота отступает (максимум на 60-70 см), а следом понижается уровень надмерзлотных вод, освобождающих, наконец, нижние - зимовочные - камеры. Вся жизнь в теплое время года сосредоточена в верхней части моховой кочки. Пунктирной линией обозначена граница между торфом и минеральным грунтом.Несправедливо не назвать безусловного рекордсмена среди лидеров - L.muscorum. В бассейне Колымы его нет, он найден в соседнем регионе - на северо-востоке Якутии, причем исключительно на участках реликтовых степей. Зимовочные камеры его гнезд, как у L.acervorum, неглубоки - 5-10 см. Но северо-восток Якутии холоднее верховьев Колымы. Здесь, вблизи полюса холода, средние абсолютные минимумы температуры в декабре-январе опускаются до –61…–63°С. Летом крошечные участки реликтовых степей не только самые теплые (максимальная температура на глубине 10 см порядка 27-29°С) среди всех других местообитаний в регионе. Они к тому же самые сухие - влажность почвы около 2-5%. Таким образом, L.muscorum приспособлен и к зимнему холоду, и к летней жаре, и к сухости субстрата. Однако эти феноменальные способности сочетаются с какой-то непонятной пока адаптивной ущербностью: вид отсутствует за пределами степных участков, что-то сдерживает его распространение. Ясно, что не потребность в высокой температуре летом, поскольку в Европе и Америке L.muscorum - несомненный широкий мезофил. Может быть, жизнь в пекле всего лишь “защита” от хищников и от конкуренции со стороны L.acervorum? Да и muscorum ли он? Нужно генетическое подтверждение…
Самый крупный муравей бореальных лесов - древоточец Camponotus herculeanus и один из самых мелких - Leptothorax acervorum. Эти муравьи - такие же атрибуты Гипоарктики, как шикша, багульники, карликовые березки и др.
Аутсайдеры
Пять других видов муравьев (четыре из рода Formica и один - Myrmica) встречаются на Верхней Колыме много реже видов-лидеров, но все же обычны настолько, что могут быть обнаружены практически на любом отрезке ее долины или на прилегающих территориях. Требования к условиям постройки гнезд и размещения зимовочных камер трех из них резко различны.
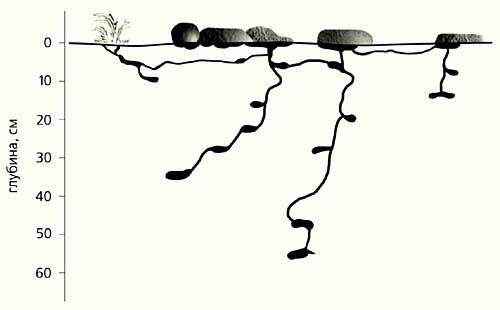
Для F.lemani главное - возможность построить гнездо с зимовочными камерами на большой глубине, где температуры будут не ниже –14…–17°С. Участок может быть даже жарким (например, на крутом степном склоне), пусть и совсем сухим (влажность поверхностных горизонтов почвы практически… отсутствует). Ходы в рыхло уложенном сланцевом плитняке шахтным стволом уходят на глубину до двух метров, где в зрелых гнездах обнаруживается сложная и протяженная сеть: там влажнее и, возможно, из почвенного воздуха конденсируется влага, достаточная для жизни. Можно выбрать и нужную для личинок температуру в градиенте “60°С на поверхности - 0°С на 2.5-3.5 м”. Чем по устройству не гнездо пустынных муравьев с его глубинными ходами для сбора влаги?! Но найдите-ка на Колыме такие участки, они очень редки…
Варианты (по степени сложности) гнезд F.lemani в зависимости от грунта и местообитания. В континентальных районах они всегда располагаются на хорошо дренируемых участках с “сухой” мерзлотой, проявляющейся только отрицательными температурами на глубине 2.5-3.5 м, да отдельными кристалликами льда, не способными скрепить грунт в ледовый бетон. Влажность почвы и грунтов вблизи поверхности ничтожна, но на глубине влага конденсируется из-за низкой температуры, поэтому гнезда F.lemani аналогичны по устройству гнездам пустынных муравьев.Муравей F.exsecta также привередлив к зимним условиям (не переносит температуры ниже –14°С), но может довольствоваться скудным летним теплом и строить гнезда чуть ли не в любых по увлажнению биотопах и располагать зимовочные камеры на глубине от 40 см до почти 2 м; под снежными надувами зимовочные камеры могут быть и в 10 см от поверхности.F.picea на юге Сибири живет почти везде: от болот и пойм до сухих степей, избегает лишь сомкнутых лесов и горных тундр. На северо-востоке же использует единственный вариант из многих возможных в других частях ареала - заселяет почти исключительно песчано-галечниковые поймы рек. Здесь он вынужден переносить длительное затопление паводками; вопреки им успевает, как и все его сородичи, развиться за один сезон от яйца до яйца (“расплод” никогда не зимует). В этих экзотических для муравьев условиях бок о бок с F.picea живет вид другого рода - M.bicolor, устраивающий подобные же гнезда, - почти полная конвергенция! Но расплод, как и у всех муравьев этого рода, зимует, а это уже иная адаптивная стратегия, и ее мы обсудим чуть позже. В поймах даже крупных рек зимой у поверхности не теплее, чем на внепойменных территориях, поэтому зимовочные камеры всегда глубоко в гальке с песком. Явно, что с поймами эти виды связывает отнюдь не смягченный вариант зимовки.
Гнездо F.picea. Его “архитектура” несложна, по устройству оно неотличимо от гнезд M.bicolor. Прогревочные камеры всегда расположены под плоской галькой, а многочисленные выходы открываются во многих местах, чаще - на голой поверхности или в куртинках чахлых растений, но никогда - в плотной тени деревьев. Ходы и камеры на глубине используются не только для зимовки, они служат убежищами во время паводков.Завершая описание устройства гнезд, заметим, что и у лидеров, и у аутсайдеров гнезда, конечно, в разной мере варьируют по расположению, используемому материалу и т.д., но их “строительный стереотип” всегда строго выдерживается. Не бывает гнезд лидеров с глубоким расположением камер и, напротив, - гнезд аутсайдеров (кроме F.exsecta) с поверхностным расположением даже в местах, где температуры зимой достаточны для успешной зимовки.“Резерв холодоустойчивости” как критерий приспособленности
Муравьи, все без исключения, относятся к организмам, избегающим замерзания и способным зимовать в состоянии переохлаждения - замерзание жидкостей тела для них губительно. Средние температуры максимального переохлаждения составляют –40°С у L.acervorum, L.muscorum, C.herculeanus; –30°С у F.gagatoides, M.kamtschatica и M.bicolor; –25°С у F.picea; –22°С у F.lemani и F.exsecta; –18°С у F.sanguinea. Максимальная зарегистрированная температура переохлаждения –58.1° (личинки L.muscorum), и это значение можно рассматривать в качестве рекордного для муравьев вообще. Порог же длительно переносимых температур у всех видов лежит выше средних величин температур переохлаждения примерно на 3-7°С.
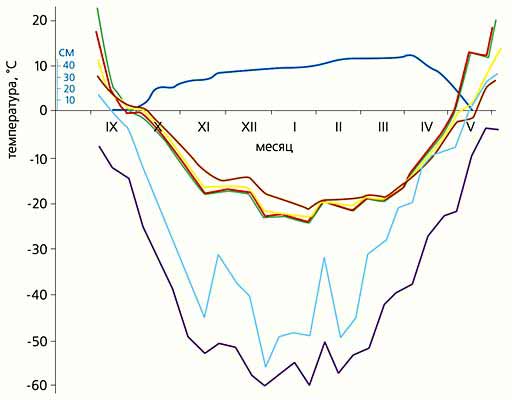
Температурные условия зимой в гнезде L.muscorum в Оймяконской котловине. Из-за раннего формирования снежного покрова (его изменение показано синей кривой) условия, приведенные на рисунке, не самые жесткие; в малоснежный год бывает много хуже, но холодоустойчивость этого вида столь велика, что погода практически не влияет на благополучие зимовки. Среднедекадные температуры почвы (на глубине 1 см - зеленая кривая, 5 см - красная, 15 см - желтая, 40 см - коричневая) и воздуха (среднесуточная - голубая кривая, абсолютный минимум за декаду - фиолетовая).В других, климатически более мягких регионах все эти показатели много выше. К примеру, на юге и севере Финляндии и даже на побережье Охотского моря под Магаданом средняя зимняя холодоустойчивость каждого изученного вида рода Formica весьма точно совпадает с ее летним значением, которую мы называем неспецифической в силу отсутствия адаптивного смысла. От лета к зиме на упомянутых территориях холодоустойчивость не растет, имеющейся неспецифической оказывается достаточно для успешной зимовки [5]. Но подобные температурные условия и еще более мягкие свойственны большей части зимней России, кроме регионов с ультраконтинентальным климатом, определяемым ядром Сибирского антициклона. В связи с этим есть основания полагать, что муравьи обходятся летней холодоустойчивостью в огромном диапазоне зимних условий, благодаря чему и была ими колонизована большая часть площади их ареалов на Севере. Лишь в значительно более жестком климате бассейна Колымы (как и других регионов, лежащих в области Якутского отрога Сибирского антициклона) муравьи должны иметь много более высокую холодоустойчивость. Насколько?Присутствие вида на данной территории само по себе означает, что вид адаптирован к местным условиям. Мерилом же степени соответствия холодоустойчивости насекомых условиям зимовки может быть “резерв холодоустойчивости”, определяемый нами как разница между порогами переносимых насекомыми температур и минимальными температурами в гнездах. Он особенно велик у L.muscorum, L.acervorum и C.herculeanus (10-15°С), что и гарантирует сохранность популяции этих видов практически повсеместно и всегда - даже в малоснежные и холодные зимы. У F.gagatoides, F.picea, M.kamtschatica и M.bicolor резерв вдвое меньше: 5-7°C. Два последних распространены предельно узко, но ограничивающие их факторы связаны с летними условиями. Полярному муравью он достаточен для крайне широкого распространения: и географического - в Гипоарктике, и биотопического - в любом, даже самом холодном районе. Более того, он может “позволить себе” строить неглубокие гнезда. По степени адаптированности к зимнему холоду полярный муравей весьма близок к рекордсменам холодоустойчивости - C.herculeanus и двум видам рода Leptothorax. В этом отношении (а не только из-за распространения) F.gagatoides оправдывает свое русское название - “полярный муравей”.
Остальные четыре вида рода Formica минимально холодостойки, резерв не превышает 2-3°С, а в неблагоприятные годы отсутствует вовсе; соответственно, и распространены они локально.
Криопротекторы и адаптивная стратегия
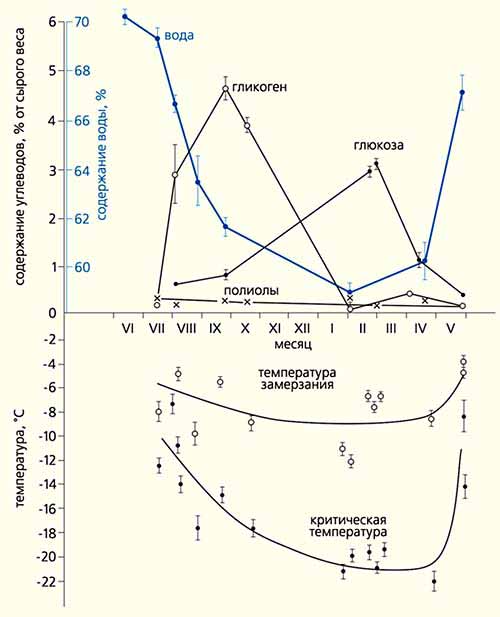
По биохимическим механизмам обеспечения холодоустойчивости и сезонной изменчивости температурных характеристик исследованные виды подразделяются на две группы. У первой группы (L.acervorum, L.muscorum, C.herculeanus, M.kamtschatica, M.bicolor) температуры максимального переохлаждения от лета к зиме меняются значительно, максимально на 25-30°С (главным образом за счет снижения температуры замерзания); в основе механизма криопротекции лежит накопление полиолов (многоатомных спиртов) до 10-20% от веса тела. Во второй группе (Formica) изменение холодоустойчивости от лета к зиме составляет только 6-12°С, температура замерзания падает лишь на 1-6° С; полиолов немного (0.4-2%), но зимой растет концентрация сахаров (3-6%).
Содержание углеводов и воды (вверху) у рабочих особей F.exsecta существенно меняется от лета к зиме, обеспечивая лишь невысокую холодоустойчивость этого вида. На температурном поле кружочками со штрихами обозначены средние значения и дисперсия показателя в разных гнездах.Таким образом, биохимические основы защиты от холода в основных чертах задаются родовым уровнем. Однако принадлежность к роду и даже подроду, по-видимому, определяет адаптивную стратегию холодоустойчивости конкретного вида далеко не полностью. Три вида - F.lemani, F.picea и F.gagatoides, относящиеся к одному подроду (Serviformica) и обитающие здесь в нехарактерных в целом для рода условиях [6], - имеют, между тем, крайне различную холодоустойчивость. У F.lemani, F.picea она весьма “средняя” (по отношению ко всему диапазону холодоустойчивости изученных видов); у F.gagatoides достигает столь значительных величин, что оказывается близкой к таковой у представителей рода Myrmica, обладающих иной биохимической обеспеченностью.В целом же наши муравьи используют три адаптивных стратегии к низким температурам зимовки. У С.herculeanus, L.acervorum и L.muscorum все построено на физиолого-биохимических возможностях - очень высокая устойчивость к холоду обеспечивается накоплением антифризов, что позволяет им зимовать непосредственно у поверхности почвы. В основе адаптивного пути второй группы (F.gagatoides, M.kamtschatica и M.bicolor) - компенсация недостаточности физиолого-биохимических возможностей поведенческими адаптациями. Эти муравьи могут жить где угодно, устраивая зимовочные камеры поглубже. Третья группа, включающая наиболее чувствительные к холоду виды из рода Formica, существует лишь в особых, благоприятных для них условиях, складывающихся в небольшой группе местообитаний; ее физиолого-биохимические возможности невелики. Таким образом, хотя физиолого-биохимическая основа задается на уровне рода, адаптивная стратегия видов сугубо индивидуальна и осуществляется различными сочетаниями физиолого-биохимических и поведенческих механизмов *. В ряде случаев интегральные адаптивные возможности представителей разных родов могут быть практически одинаковы.
* На юге и севере Финляндии и даже на побережье Охотского моря под Магаданом средняя зимняя холодоустойчивость каждого изученного вида рода Formica весьма точно совпадает с ее летним значением. От лета к зиме холодоустойчивость не растет - муравьи обходятся летней. Лишь в жестком климате регионов, лежащих в области Якутского отрога Сибирского антициклона, муравьи должны иметь много более высокую устойчивость к холоду.Ярчайший пример тому - ключевой для понимания адаптивной стратегии вид - полярный муравей. Ко многому уже сказанному о нем добавим, что его изначальный адаптивный потенциал принципиально ограничен по сравнению с близкими с ним по холодоустойчивости L.acervorum и C.herculeanus. Дело в том, что они способны растягивать онтогенез на несколько лет за счет зимовки личинок; полярный же муравей - “однолетник”, а это значит, что он нуждается для прохождения онтогенеза в повышенном и одновременно гарантированном количестве тепла (если тепла будет недостаточно, расплод погибнет). В целом же эффект использования разных стратегий всех этих муравьев, оцениваемый, например, по численности гнезд и широте спектра населяемых биотопов, оказывается также весьма близок.Холодоустойчивость и распространение
В какой мере реализован потенциал холодоустойчивости у муравьев верховий Колымы? Мы полагаем, что полученные характеристики для большинства видов, по-видимому, близки к предельно возможным для всех муравьев. Косвенно об этом свидетельствует их географическое распространение.
На запад от верховий Колымы к верховьям Индигирки идет постепенное выпадение всех видов рода Formica, кроме F.gagatoides, F.picea и, возможно, F.fusca (см. табл.). Поскольку сравниваемые фауны относятся к регионам, удаленным от океана и лежащим на одной широте, постольку дело не в летнем тепле, а только в зимних условиях. На Колыме резерв холодоустойчивости для рода Formica (повторяем, кроме полярного муравья и F.picea) очень мал - всего 2-3°С. В Оймяконской же котловине средняя многолетняя январская температура воздуха (по данным одноименной метеостанции) составляет –49.5°С (при среднемноголетнем минимуме –64°С) против –35.2°С в верховьях Колымы (метеостанция Усть-Омчуг). Мощность снежного покрова в самые холодные месяцы на Колыме в полтора-два раза больше, чем на Индигирке (40-50 и 30 см соответственно). Как следствие, среднемесячные температуры почвы на глубине 20-40 см в верхней части бассейна Индигирки падают до –23…-24°С против –15…–18°С в верховьях Колымы. Понятно, что резерва холодоустойчивости у муравьев рода Formica (за названными исключениями) на Индигирке в таких условиях не остается, поэтому и самих видов нет.
Еще западнее - в Центральной Якутии - фауна муравьев содержит не менее 24 видов [7], т.е. богаче фауны бассейна Колымы на 10 видов, а бассейна Индигирки - на 14. Муравьев рода Formica там 14 видов против 9 в Магаданской обл. (см. табл.). Очевидно, что дело в климате: в Якутске и летом, и, главное, зимой теплее, чем в Оймяконе, - почти на 6°С выше средняя температура воздуха в январе, а минимумы в почве на глубине 20-40 см не превосходят –15…–18°С, что даже выше, чем в верховьях Колымы.
А вот распространение многих видов на север и северо-восток, вероятно, ограничивают иные факторы. Это нетрудно показать. Большинство видов муравьев севернее границы леса не идет. Только по сухим и теплым бровкам речных террас и морен в горных тундрах, выдвинутых в тундровую зону, а также в кустарниковых тундрах встречаются полноценные гнезда L.acervorum. Завершение этой тенденции проявляется в отсутствии каких бы то ни было муравьев в зональных тундрах. На Аляске ситуация аналогична. Между тем, тундры Чукотки лежат в области январских изотерм от –20°С (метеостанция Уэлен) до –28°С (метеостанция Певек); мощности снежного покрова в верховьях р.Колымы и на Чукотке близки. Поэтому зимние температуры почв в тундрах даже в среднем не ниже таковых в верховьях Колымы и, тем более, Индигирки. Поскольку C.herculeanus, L.acervorum и F.gagatoides широко распространены в верховьях Колымы, Индигирки и Яны, повторимся - на территориях с самой суровой зимой из возможных в Северном полушарии, можно считать отсутствие этих муравьев в тундровой зоне связанным с летними обстоятельствами.
Если зима еще суровее...
Вот так и зимуют муравьи на Северо-Востоке; добавим - сейчас, в конкретной климатической обстановке. Однако она, как известно было еще Кузьме Пруткову (“…всего же непостояннее петербургская атмосфера”), переменчива даже на коротких временах, не говоря уже о том, что каких-то 10-12 тыс. лет назад был плейстоцен с его ледниковыми катаклизмами. Как сказывается на благополучии муравьев ужесточение зимы?
Понятно, что виды с большим резервом холодоустойчивости безразличны к понижению температур на глубине расположения зимовочных камер. Более того, они явно благоденствуют. Нетрудно предположить, что благодаря снижению общего биоразнообразия, ослаблению пресса хищников и конкурентов здесь, на краю ареала, складывается “биотический оптимум” [8]. Именно с этим, скорее всего, связана уже упоминавшаяся колоссальная численность L. acervorum в нашем регионе.
Не менее показательный пример - удивительный ареал полярного муравья [9], лежащий к северу от 60° с.ш. (хотя, наверное, по горным системам этот вид может проникать вместе с охотской биотой до бассейна Амура). Понятно, что южнее 60°с.ш. гидротермические условия для полярного муравья не хуже. Сюда его не пускают, вероятнее всего, свои же собратья - весьма обширная группа лесных видов рода Formica из числа широких палеарктов.
Свидетельством обстановки с ослабленной конкуренцией, складывающейся в горно-лесных районах Северо-Восточной Азии, служит крайне слабая видовая насыщенность комплексов муравьев. Число совместно обитающих видов обычно не превосходит четырех; лишь в единственном из многих десятков изученных биотопов к гипоарктической троице присоединяются F.exsecta и F.sanguinea. Обычно же добавляется только один из аутсайдеров.
Выносливость гипоарктического комплекса муравьев поразительна. Мало того, что они вездесущи ныне. Остатки хитина C.herculeanus, L.acervorum и двух видов рода Formica найдены в отложениях позднего плейстоцена ряда разрезов северо-востока Азии [10]. Один из них, несомненно, F.gagatoides, второй, вероятнее всего, F.picea. Не содержится ли в этих фактах указание на еще более высокую холодоустойчивость муравьев, нежели выявлена нами? Если нет, то ископаемые остатки муравьев можно использовать в качестве надежных температурных индикаторов, в соответствии с которыми придется корректировать наши представления о зимних условиях плейстоцена.
Одним словом, нужно перепроверить предельные значения холодоустойчивости муравьев. А для этого нужна экспедиция в Северо-Восточную Якутию, в район полюса холода - все-таки поближе к… плейстоцену.
Работа выполнена при поддержке Российского фонда фундаментальных исследований. Проект 04-04-48187.

Снежная зима в Северо-Восточной Якутии.
В центре антициклона ветер - крайняя редкость, и снежная кухта может лежать на деревьях до весны.
Литература
1. Ушатинская Р.С. Скрытая жизнь и анабиоз. М., 1990.
2. Insects at low temperature / Eds R.E.Lee, D.L.Delinger. N.Y., 1991. P.1-513.
3. Хансен Т.Э., Вийк М.О. // Зоол. журн. 1981. Т.60. Вып.3. С.380-387.
4. Купянская А.Н. Муравьи (Hymenoptera, Formicidae) Дальнего Востока СССР. Владивосток, 1990.
5. Берман Д.И., Жигульская З.А. // Успехи современной биологии. 1995. Т.115. Вып.6. С.677-691.
6. Чернов Ю.И. // Зоол. журн. 1988. Т.67. Вып.10. С.1445-1457.
7. Дмитриенко В.К., Петренко Е.С. Муравьи таежных биоценозов Сибири. Новосибирск, 1976.
8. Арнольди К.В. // Зоол. журн. 1957. Т.36. Вып.11. С.1609-1629.
9. Длусский Г.М. Муравьи рода формика. М., 1967.
10. Киселев С.В. Позднекайнозойские жесткокрылые северо-востока Сибири. М., 1981.